Última actualizacón: 15 mayo 2020 a las 16:16


Dos modelos acerca de la colonización de América
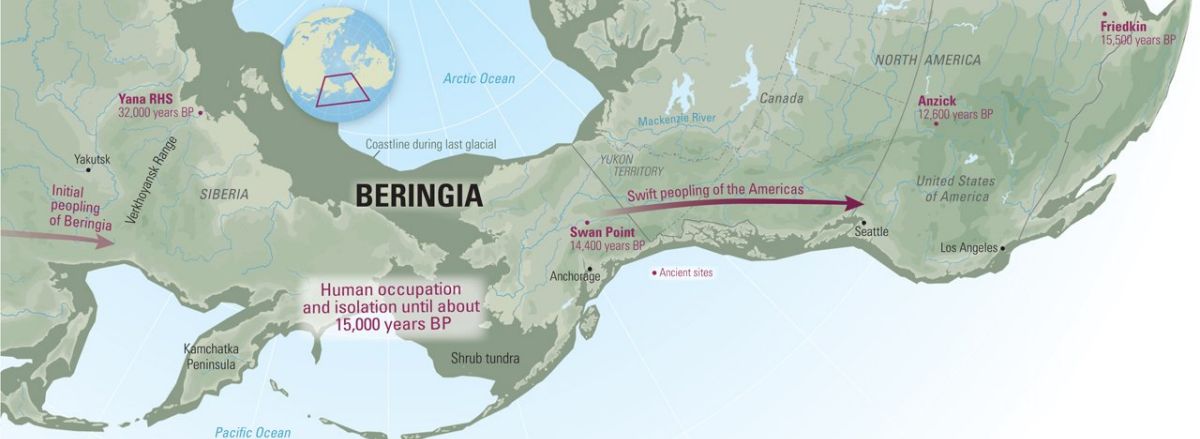
El modelo del «poblamiento rápido»
Como indicamos en la primera parte de esta serie de anotaciones, los investigadores mantienen dos posturas acerca de la función que cumplió Beringia en lo que hace referencia al poblamiento de América. Algunos sostienen que las poblaciones que migraron hacia América se detuvieron en la región durante miles de años, dejando una impronta genética en los nativos americanos. Es decir, Beringia pudo haber sido un lugar más cómodo para vivir durante el UMG de lo que se pensaba hasta hace poco, actuando como un «refugio» ante el duro clima circundante.
Sin embargo, otros defienden que Beringia no fue más que un mero «pasillo», un «puente» que tanto los grandes mamíferos como nuestros antepasados se apresuraron a cruzar en su camino desde Asia al nuevo continente: se trata de la llamada hipótesis del «poblamiento rápido», y según este modelo el aislamiento se habría producido en Asia y no en la masa terrestre de Beringia.
Su argumentación es la siguiente: dado que desde hace 40000 años las masas de hielo de los glaciares cubrían gran parte de lo que hoy es Alaska y Canadá –impidiendo el paso hacia América 1–, los primeros pobladores del continente hubieron de esperar hasta que se formaron unos corredores tras la retirada del hielo, bien por la costa (hace unos 15000 años), o bien por el interior (hace entre 14000 y 13500 años).
Analicemos los datos que apoyan esta versión.
Datos arqueológicos
Las herramientas de yacimientos como Swan Point, en el centro de Alaska, datado en 14000 años de antigüedad; el de Nenana, con una antigüedad de entre 13800 y 13000 años; y el de Ushki en Kamchatka (13000 años), constituyen las pruebas arqueológicas más tempranas y fiables que se han hallado hasta el momento en el este de Beringia.
Por otro lado, hemos de tener en cuenta que el registro arqueológico de los corredores que quedaron libres de hielo ya dentro del continente americano, aunque mejora con los recientes estudios y a pesar de que hay resultados prometedores, todavía es insuficiente para resolver algunas de las dudas que quedan pendientes en lo tocante a la presencia de poblaciones humanas.
Por lo tanto, los defensores del «poblamiento rápido» se apoyan en la ausencia de pruebas arqueológicas, en la falta de un enlace claro entre el yacimiento siberiano de Yana RHS (que analizamos en la segunda parte de esta serie) y Swan Point 2.

El no haber encontrado yacimientos más antiguos hace suponer que las duras condiciones climáticas impidieron el poblamiento de América antes de que el deshielo de los glaciares hace aproximadamente 14000 años, permitiera el paso seguro. Los yacimientos de Monte Verde en Chile, Schaefer y Hebior, apuntan a esa época como la de la primera presencia humana en América. Estos yacimientos, junto a los de Meadowcroft, Page-Ladson y la cueva de Paisley, podrían representar el estrato más antiguo de la prehistoria americana y, por tanto, el origen de la cultura Clovis.
Si Beringia no fue un «refugio», ¿dónde se detuvieron las poblaciones en su camino a América mientras mejoraban las condiciones climáticas?
Dos investigadores 3 han analizado la tecnología de microláminas de diferentes yacimientos del noreste de Asia y de Beringia para concluir que la situación climática durante el UMG llevó a las poblaciones hacia las islas de Sajalín y Hokkaido. Por lo tanto, defienden que la migración desde el interior de Siberia se produjo en primer lugar hacia sur y el este, no directamente hacia Beringia.
Durante el UMG ambas islas estaban unidas entre sí y, al mismo tiempo, con tierra firme. Desde hace 26000 años, los signos de habitación humana en esta región aumentan bruscamente, lo que demostraría la existencia de un flujo más o menos constante de llegada de personas. Y esto sucede poco después de que el número de yacimientos humanos en el sur de Siberia descienda bruscamente, como si la gente se estuviera desplazando de un lugar a otro. Para ellos, el «refugio» no estuvo en Beringia sino en la península formada por esas islas.
Información genética
El trabajo de Raghavan et al. (2015) que analizamos en la anterior anotación planteaba que las duras condiciones climáticas habían provocado el aislamiento geográfico de los antepasados de los nativos americanos, que conllevó a su vez el aislamiento genético. Ofrecieron como fecha más probable para esta separación la de 22000 años, aunque parece que el flujo genético continuó hasta hace unos 12000 años, cuando el puente de Beringia quedó sumergido bajo el mar. Respecto a la cuestión de si ese aislamiento se había producido en Siberia o en Beringia quedó sin respuesta.
Los análisis completos tanto de ADN moderno como antiguo indican que las poblaciones de nativos americanos divergieron de sus antepasados asiáticos hace no más de 23000 años, demasiado poco tiempo para que tuviera lugar el aislamiento genético defendido por la «hipótesis de la parada en Beringia».
Por otro lado, recientes estudios de ADN 4 sugieren que esa divergencia se produjo mucho antes, hace 36000 años –con una separación definitiva hace entre 25000 y 20000 años–, lo que supondría que la separación geográfica entre las poblaciones se produjo mucho antes de lo que sugiere la «hipótesis de la parada en Beringia».
En cualquier caso debemos ser cautos con este tipo de estudios genéticos: el cálculo del reloj molecular puede plantear problemas metodológicos, es decir, la estimación de la tasa de mutación que sirve para establecer fechas aún no es una cuestión zanjada. Por ello, pese al hallazgo en el ártico canadiense (yacimiento de Bluefish) de marcas de corte en huesos con una antigüedad de 23700 años, y que podría documentar la presencia humana en el este de Beringia, el hecho de que sea un yacimiento aislado y que no se hayan recuperado herramientas de piedra o pruebas del uso del fuego plantean dudas acerca de si esos yacimientos demuestran realmente la presencia humana continuada en la región.
Volviendo al patrón genético que vemos en los múltiples estudios que se están realizando, puede haber una explicación alternativa: el «surfing»genético. Se trata de un cambio genético acelerado que se produce cuando una población pequeña se extiende muy rápido por un área geográfica muy grande 5. Esta podría ser una explicación alternativa a la que han ofrecido los defensores de la parada en Beringia.
Paleoecología
Varios estudios apuntan a que la migración hacia América fue viable hace unos 18000 años, al confirmar que el margen occidental de la capa de hielo que constituía el Glaciar de la Cordillera se había retirado antes de lo que se pensaba. Este temprano deshielo dejó expuestas numerosas islas que podrían haber sido utilizadas por las primeras personas que emigraron hacia el sur dentro del continente 6.
En 2016 7 se publicaron los primeros resultados del análisis de un enorme yacimiento SITUADO en el Ártico central de Siberia, datado en alrededor de 48000 años. Se encontraron los huesos de un mamut con evidentes marcas de corte. Este yacimiento amplía el área de poblamiento hasta casi los 72° N. La capacidad de nuestros antepasados para cazar mamuts les permitió sobrevivir y extenderse ampliamente por la Siberia ártica más septentrional y supuso un cambio cultural y adaptativo notable. Y lo que es más importante, la presencia de Homo sapiens en esta zona cercana al puente de Beringia pudo haberles brindado la oportunidad de entrar en el Nuevo Mundo antes del UMG.

En cualquier caso, como hemos venido comentado, el mayor problema a la hora de establecer cuándo y cómo se produjo la entrada en América es la ausencia de pruebas físicas directas de la ocupación humana de Beringia que permitan poner a prueba las interpretaciones de los datos genéticos.
Conclusiones
Con todos los datos sobre la mesa, lo que podemos afirmar con seguridad es que la colonización de Beringia no se produjo al mismo tiempo que el poblamiento de América. Mucho antes de la entrada de los humanos en el nuevo continente se asentaron en la parte norte de Eurasia. Contamos con bastantes pruebas de que Homo sapiens colonizó la región hace entre 50000 y 40000 años.
Sin embargo la hipótesis de la «parada en Beringia» sólo podrá confirmarse cuando se encuentren fósiles humanos o pruebas arqueológicas fehacientes de la presencia de poblaciones en Beringia durante el UMG y pruebas que vinculen esas poblaciones con uno o varios de los linajes de Nativos Americanos que se dispersaron a lo largo del hemisferio occidental después de hace 15000 años (cuando terminó el UMG).
Por otro lado, unir el registro arqueológico con los datos genéticos es complicado. Por ejemplo, los movimientos a gran escala de las personas y sus genes pueden producirse de diferentes formas. Además de «colonizar» una región que estuviera desocupada con anterioridad, los migrantes también pudieron acceder a regiones que ya estaban ocupadas por otras poblaciones y, gradualmente, relacionarse con ellos. Pero también existe la posibilidad de que se produjera un completo reemplazo físico de los habitantes preexistentes.
Como estamos acostumbrados a decir en este blog, serán necesarios más hallazgos, nuevas investigaciones, la aplicación de métodos novedosos, para poder despejar las dudas que envuelven a un tema de tanta importancia como fue la forma en que nuestros antepasados llegaron al continente americano.
Esperamos contarlos en esta bitácora.
Referencias
Bond, J. D. (2019), Paleodrainage map of Beringia. Yukon Geological Survey. Open File 2019-2.
Buvit, I. y Terry, K. (2016), «Outside Beringia: Why the northeast asian Upper Paleolithic record does not support a long standstill model«. PaleoAmerica, vol. 2, núm. 4, p. 281-285.
Darvill, C. M., et al. (2018), «Retreat of the western Cordilleran ice sheet margin during the last deglaciation«. Geophysical Research Letters, vol. 45, núm. 18, p. 9710-9720.
Goebel, T.; Waters, M. R. y O’Rourke, D. H. (2008), «The Late Pleistocene dispersal of modern humans in the Americas«. Science, vol. 319, núm. 5869, p. 1497-1502.
Hoffecker, J. F., et al. (2016), «Beringia and the global dispersal of modern humans«. Evolutionary Anthropology: Issues, News, and Reviews, vol. 25, núm. 2, p. 64-78.
Hoffecker, J. F.; Elias, S. A. y O’Rourke, D. H. (2014), «Out of Beringia?». Science, vol. 343, núm. 6174, p. 979-980.
Lesnek, A. J., et al. (2018), «Deglaciation of the Pacific coastal corridor directly preceded the human colonization of the Americas«. Science Advances, vol. 4, núm. 5, p.
Llamas, B., et al. (2016), «Ancient mitochondrial DNA provides high-resolution time scale of the peopling of the Americas«. Science Advances, vol. 2, núm. 4, p. e1501385.
Meiri, M., et al. (2014), «Faunal record identifies Bering isthmus conditions as constraint to end-Pleistocene migration to the New World«. Proceedings of the Royal Society B: Biological Sciences, vol. 281, núm. 1776, p. 20132167.
Moreno-Mayar, J. V., et al. (2018), «Terminal Pleistocene Alaskan genome reveals first founding population of Native Americans«. Nature, vol. 553, núm. 7687, p. 203-207.
Pitulko, V. V., et al. (2004), «The Yana RHS Site: humans in the arctic before the Last Glacial Maximum«. Science, vol. 303, núm. 5654, p. 52-56.
Pitulko, V. V., et al. (2012), «The oldest art of the Eurasian Arctic: personal ornaments and symbolic objects from Yana RHS, Arctic Siberia«. Antiquity, vol. 86, núm. 333, p. 642-659.
Pitulko, V. V., et al. (2016), «Early human presence in the Arctic: Evidence from 45,000-year-old mammoth remains«. Science, vol. 351, núm. 6270, p. 260-263.
Pitulko, V.; Pavlova, E. y Nikolskiy, P. (2017), «Revising the archaeological record of the Upper Pleistocene arctic Siberia: Human dispersal and adaptations in MIS 3 and 2«. Quaternary Science Reviews, vol. 165, p. 127-148.
Pringle, H. (2014), «Welcome to Beringia«. Science, vol. 343, núm. 6174, p. 961-963.
Raghavan, M., et al. (2015), «Genomic evidence for the Pleistocene and recent population history of Native Americans«. Science, vol. 349, núm. 6250, p. aab3884.
Tamm, E., et al. (2007), «Beringian standstill and spread of Native American founders«. PLoS ONE, vol. 2, núm. 9, p. e829.
Watson, T. (2017), «News Feature: Is theory about peopling of the Americas a bridge too far?«. Proceedings of the National Academy of Sciences, vol. 114, núm. 22, p. 5554-5557.
Bibliografía recomendada
Goebel, T. y Buvit, I. (2011), From the Yenisei to the Yukon. Interpreting lithic assemblage variability in Late Pleistocene-Early Holocene Beringia. College Station: Texas A&M University Press, 416 p.
West, F. H. y West, C. F. (1996), American beginnings. The prehistory and palaeoecology of Beringia. Chicago: University of Chicago Press, xxi, 576 p.

Notas
- Quienes defienden esta hipótesis ven cualquier afirmación de una colonización de América antes de esa fecha con mucho escepticismo. ↩
- Aunque, por el contrario, se argumenta que ese vacío de 16000 años en el registro arqueológico se debe a que los yacimientos que podrían arrojar luz sobre esta cuestión están bajo el agua. ↩
- Buvit y Terry (2016). ↩
- Como el de Moreno-Mayar et al. (2018). ↩
- Un ejemplo de este proceso se ha analizado en relación a la tortuga mora en Murcia. ↩
- Ver, en este sentido, Lesnek et al. (2018) y Darvill et al. (2018). ↩
- Pitulko et al (2016). ↩














