Título: Disclosing the past. An autobiography Autor: Mary Leakey Edita: Weidenfeld & Nicolson, 1984 Encuadernación: Tapa dura. Número de páginas: 224 p. ISBN: 0297785451
Reseña del editor
La reconocida arqueóloga ofrece una mirada incisiva y detallada de su notable familia y analiza su trabajo con su marido en África Oriental, así como sus descubrimientos, que alteraron para siempre el curso de la antropología moderna.
Reseña
Lo primero que debemos señalar en honor a la verdad es que no estamos ante una verdadera «autobiografía». Mary Leakey, siempre reacia a hablar de su vida privada, necesitó de la ayuda de un escritor profesional para dar forma al texto que ahora tenemos entre manos. En cualquier caso, este dato no desmerece el contenido ni la importancia de contar con un libro gracias al que podemos conocer de primera mano las inquietudes y vivencias de quien ha sido una de las arqueólogas más importantes del siglo XX.
Lo segundo, pese a que a estas alturas no debiera ser necesario, es apuntar que Mary Leakey no fue solo «la mujer de Louis Leakey». Aún hay textos en los que se deja entrever que ese dato es todo lo que tenemos que saber. En realidad, la valía profesional e intelectual, así como los logros científicos de Mary Leakey superaron en muchos aspectos a los de su marido.
Mary divide su biografía en tres fases: desde la niñez hasta que conoce a Louis Leakey; sus años con él que terminan con al morir en 1972 y, por último, el periodo «post-Louis», donde destacan sus hallazgos en Laetoli.
Sin embargo, ella reconoce que todo comenzó en el verano de 1935 cuando, con apenas 22 años, visitó por primera vez África Oriental. Tras conducir por la ladera rocosa del cráter del Ngorongoro en Tanzania, se encontró frente a una vista que dejó una huella indeleble en su espíritu y marcó su futuro. Esa vista era la de la vasta llanura del Serengeti, «que se extendía hasta el horizonte como el mar… siempre igual, pero siempre diferente».
Según sus palabras, ese fue el momento en que África se apoderó de ella. A pesar de que vivió y viajó por diferentes lugares del mundo, los barrancos, los cauces de los arroyos, los acantilados y las laderas de ese paisaje africano iban a constituir su hogar durante casi medio siglo. Fue allí donde se vio involucrada en algunos de los hallazgos más importantes y dramáticos que han rodeado al mundo de la arqueología prehistórica y el estudio de la evolución humana.
Mary Nicol nació en Londres en 1913, hija única de Cecilia Frere y Erskine Nicol, un pintor de éxito de quien heredó no solo el amor al aire libre y la curiosidad por la vida de nuestros antepasados, sino un destacado talento para el dibujo. Era descendiente de John Frere, un anticuario (como se los conocía entonces) que encontró diversas herramientas de piedra y que en 1797 argumentó que fueron hechas por quienes aún no conocían el metal -una idea considerada por aquel entonces casi una herejía-.
Mary nos cuenta que tuvo una infancia alegre, dividida entre Londres y Europa, a donde su padre viajaba cada año para pintar. También nos explica que hacía sufrir terriblemente a las jóvenes institutrices que sus padres contrataban para darle una educación formal. En el suroeste de Francia comenzó su interés por la arqueología, participando en excavaciones arqueológicas y recogiendo herramientas de piedra con su padre. Por supuesto, también visitó las famosas pinturas rupestres de esa región. Tras la muerte de su padre, su madre y ella se vieron obligadas a regresar a Inglaterra, en ese momento sus intereses cambiaron hacia la historia y arqueologías británicas. Al final de su adolescencia ya conocía a muchos de los principales arqueólogos de la época y tenía perfectamente claro a qué quería dedicarse el resto de su vida. Sus visitas a Stonehenge y Avebury no hicieron sino reforzar ese interés.
Su sobresaliente capacidad para dibujar herramientas de piedra le llevó a reunirse con Louis Leakey, que buscaba un ilustrador para uno de sus libros, dando así comienzo la segunda fase de su vida.
En 1935 visitó Kenia y Tanzania con él, y en 1936 se casaron y se mudaron al este de África. Aquí profundizamos en los primeros años de sus trabajos en la garganta de Olduvai; los sempiternos problemas de financiación que obligan a Louis a centrarse en el objetivo de conseguir patrocinadores; el nacimiento de cuatro hijos (la pequeña Deborah murió a los tres meses de disentería) y el descubrimiento de Zinjanthropus boisei.
La muerte de Louis obligó a Mary a ocupar el centro del «escenario». Además de dirigir sus proyectos de investigación, tuvo que asumir los papeles de recaudadora de fondos, organizadora, publicista y conferenciante (roles que Louis había desempeñado siempre con enorme energía y entusiasmo). Para ella supuso un enorme desafío ya que se desenvolvía con más soltura en un yacimiento que en un salón repleto de público. Sin embargo, superado el rechazo inicial, lo aceptó con determinación y pronto destacó en la escena internacional, siendo reclamada como conferenciante en todo el mundo.
No vamos a encontrar en este libro referencias a la bochornosa conducta de Louis Leakey, tanto en lo personal como en lo profesional. Mary se limita a decir que llegó un momento en sus vidas en que ella había perdido el respeto por su marido, aunque refiriéndose más a su capacidad intelectual que a sus relaciones extramatrimoniales (la insistencia de Louis en defender el yacimiento californiano de Calico tuvo un papel determinante). Estuvieron bastante tiempo haciendo vidas separadas antes de su fallecimiento en 1972.
Estamos, en definitiva, ante un texto imprescindible para todo amante de la arqueología y la historia de la ciencia.
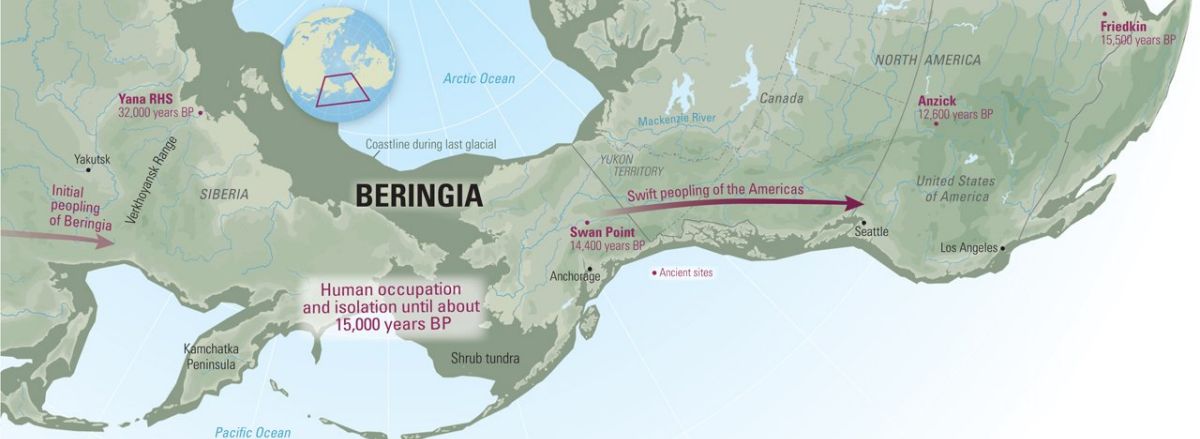
Como indicamos en la primera parte de esta serie de anotaciones, los investigadores mantienen dos posturas acerca de la función que cumplió Beringia en lo que hace referencia al poblamiento de América. Algunos sostienen que las poblaciones que migraron hacia América se detuvieron en la región durante miles de años, dejando una impronta genética en los nativos americanos. Es decir, Beringia pudo haber sido un lugar más cómodo para vivir durante el UMG de lo que se pensaba hasta hace poco, actuando como un «refugio» ante el duro clima circundante.
Sin embargo, otros defienden que Beringia no fue más que un mero «pasillo», un «puente» que tanto los grandes mamíferos como nuestros antepasados se apresuraron a cruzar en su camino desde Asia al nuevo continente: se trata de la llamada hipótesis del «poblamiento rápido», y según este modelo el aislamiento se habría producido en Asia y no en la masa terrestre de Beringia.
Su argumentación es la siguiente: dado que desde hace 40000 años las masas de hielo de los glaciares cubrían gran parte de lo que hoy es Alaska y Canadá –impidiendo el paso hacia América 1–, los primeros pobladores del continente hubieron de esperar hasta que se formaron unos corredores tras la retirada del hielo, bien por la costa (hace unos 15000 años), o bien por el interior (hace entre 14000 y 13500 años).
Analicemos los datos que apoyan esta versión.
Datos arqueológicos
Las herramientas de yacimientos como Swan Point, en el centro de Alaska, datado en 14000 años de antigüedad; el de Nenana, con una antigüedad de entre 13800 y 13000 años; y el de Ushki en Kamchatka (13000 años), constituyen las pruebas arqueológicas más tempranas y fiables que se han hallado hasta el momento en el este de Beringia.
Por otro lado, hemos de tener en cuenta que el registro arqueológico de los corredores que quedaron libres de hielo ya dentro del continente americano, aunque mejora con los recientes estudios y a pesar de que hay resultados prometedores, todavía es insuficiente para resolver algunas de las dudas que quedan pendientes en lo tocante a la presencia de poblaciones humanas.
Por lo tanto, los defensores del «poblamiento rápido» se apoyan en la ausencia de pruebas arqueológicas, en la falta de un enlace claro entre el yacimiento siberiano de Yana RHS (que analizamos en la segunda parte de esta serie) y Swan Point 2.
La combinación de datos genéticos y arqueológicos de Siberia, Beringia y de América sugieren que los humanos llegamos al continente americano desde el sur de Siberia poco después del UMG, hace unos 15000 años. Tomado de Goebel, Waters, y O’Rourke (2008).
El no haber encontrado yacimientos más antiguos hace suponer que las duras condiciones climáticas impidieron el poblamiento de América antes de que el deshielo de los glaciares hace aproximadamente 14000 años, permitiera el paso seguro. Los yacimientos de Monte Verde en Chile, Schaefer y Hebior, apuntan a esa época como la de la primera presencia humana en América. Estos yacimientos, junto a los de Meadowcroft, Page-Ladson y la cueva de Paisley, podrían representar el estrato más antiguo de la prehistoria americana y, por tanto, el origen de la cultura Clovis.
Si Beringia no fue un «refugio», ¿dónde se detuvieron las poblaciones en su camino a América mientras mejoraban las condiciones climáticas?
Dos investigadores 3 han analizado la tecnología de microláminas de diferentes yacimientos del noreste de Asia y de Beringia para concluir que la situación climática durante el UMG llevó a las poblaciones hacia las islas de Sajalín y Hokkaido. Por lo tanto, defienden que la migración desde el interior de Siberia se produjo en primer lugar hacia sur y el este, no directamente hacia Beringia.
Durante el UMG ambas islas estaban unidas entre sí y, al mismo tiempo, con tierra firme. Desde hace 26000 años, los signos de habitación humana en esta región aumentan bruscamente, lo que demostraría la existencia de un flujo más o menos constante de llegada de personas. Y esto sucede poco después de que el número de yacimientos humanos en el sur de Siberia descienda bruscamente, como si la gente se estuviera desplazando de un lugar a otro. Para ellos, el «refugio» no estuvo en Beringia sino en la península formada por esas islas.
Información genética
El trabajo de Raghavan et al. (2015) que analizamos en la anterior anotación planteaba que las duras condiciones climáticas habían provocado el aislamiento geográfico de los antepasados de los nativos americanos, que conllevó a su vez el aislamiento genético. Ofrecieron como fecha más probable para esta separación la de 22000 años, aunque parece que el flujo genético continuó hasta hace unos 12000 años, cuando el puente de Beringia quedó sumergido bajo el mar. Respecto a la cuestión de si ese aislamiento se había producido en Siberia o en Beringia quedó sin respuesta.
Los análisis completos tanto de ADN moderno como antiguo indican que las poblaciones de nativos americanos divergieron de sus antepasados asiáticos hace no más de 23000 años, demasiado poco tiempo para que tuviera lugar el aislamiento genético defendido por la «hipótesis de la parada en Beringia».
Por otro lado, recientes estudios de ADN 4 sugieren que esa divergencia se produjo mucho antes, hace 36000 años –con una separación definitiva hace entre 25000 y 20000 años–, lo que supondría que la separación geográfica entre las poblaciones se produjo mucho antes de lo que sugiere la «hipótesis de la parada en Beringia».
En cualquier caso debemos ser cautos con este tipo de estudios genéticos: el cálculo del reloj molecular puede plantear problemas metodológicos, es decir, la estimación de la tasa de mutación que sirve para establecer fechas aún no es una cuestión zanjada. Por ello, pese al hallazgo en el ártico canadiense (yacimiento de Bluefish) de marcas de corte en huesos con una antigüedad de 23700 años, y que podría documentar la presencia humana en el este de Beringia, el hecho de que sea un yacimiento aislado y que no se hayan recuperado herramientas de piedra o pruebas del uso del fuego plantean dudas acerca de si esos yacimientos demuestran realmente la presencia humana continuada en la región.
Volviendo al patrón genético que vemos en los múltiples estudios que se están realizando, puede haber una explicación alternativa: el «surfing»genético. Se trata de un cambio genético acelerado que se produce cuando una población pequeña se extiende muy rápido por un área geográfica muy grande 5. Esta podría ser una explicación alternativa a la que han ofrecido los defensores de la parada en Beringia.
Paleoecología
Varios estudios apuntan a que la migración hacia América fue viable hace unos 18000 años, al confirmar que el margen occidental de la capa de hielo que constituía el Glaciar de la Cordillera se había retirado antes de lo que se pensaba. Este temprano deshielo dejó expuestas numerosas islas que podrían haber sido utilizadas por las primeras personas que emigraron hacia el sur dentro del continente 6.
En 2016 7 se publicaron los primeros resultados del análisis de un enorme yacimiento SITUADO en el Ártico central de Siberia, datado en alrededor de 48000 años. Se encontraron los huesos de un mamut con evidentes marcas de corte. Este yacimiento amplía el área de poblamiento hasta casi los 72° N. La capacidad de nuestros antepasados para cazar mamuts les permitió sobrevivir y extenderse ampliamente por la Siberia ártica más septentrional y supuso un cambio cultural y adaptativo notable. Y lo que es más importante, la presencia de Homo sapiens en esta zona cercana al puente de Beringia pudo haberles brindado la oportunidad de entrar en el Nuevo Mundo antes del UMG.
Yacimiento SK. Los restos de un mamut con evidentes marcas de corte y de haber sido manipulado tras su muerte, demuestran la presencia humana en la región hace unos 49000 años. Tomado de Pitulko et al (2016).
En cualquier caso, como hemos venido comentado, el mayor problema a la hora de establecer cuándo y cómo se produjo la entrada en América es la ausencia de pruebas físicas directas de la ocupación humana de Beringia que permitan poner a prueba las interpretaciones de los datos genéticos.
Conclusiones
Con todos los datos sobre la mesa, lo que podemos afirmar con seguridad es que la colonización de Beringia no se produjo al mismo tiempo que el poblamiento de América. Mucho antes de la entrada de los humanos en el nuevo continente se asentaron en la parte norte de Eurasia. Contamos con bastantes pruebas de que Homo sapiens colonizó la región hace entre 50000 y 40000 años.
Sin embargo la hipótesis de la «parada en Beringia» sólo podrá confirmarse cuando se encuentren fósiles humanos o pruebas arqueológicas fehacientes de la presencia de poblaciones en Beringia durante el UMG y pruebas que vinculen esas poblaciones con uno o varios de los linajes de Nativos Americanos que se dispersaron a lo largo del hemisferio occidental después de hace 15000 años (cuando terminó el UMG).
Por otro lado, unir el registro arqueológico con los datos genéticos es complicado. Por ejemplo, los movimientos a gran escala de las personas y sus genes pueden producirse de diferentes formas. Además de «colonizar» una región que estuviera desocupada con anterioridad, los migrantes también pudieron acceder a regiones que ya estaban ocupadas por otras poblaciones y, gradualmente, relacionarse con ellos. Pero también existe la posibilidad de que se produjera un completo reemplazo físico de los habitantes preexistentes.
Como estamos acostumbrados a decir en este blog, serán necesarios más hallazgos, nuevas investigaciones, la aplicación de métodos novedosos, para poder despejar las dudas que envuelven a un tema de tanta importancia como fue la forma en que nuestros antepasados llegaron al continente americano.
Goebel, T. y Buvit, I. (2011), From the Yenisei to the Yukon. Interpreting lithic assemblage variability in Late Pleistocene-Early Holocene Beringia. College Station: Texas A&M University Press, 416 p.
West, F. H. y West, C. F. (1996), American beginnings. The prehistory and palaeoecology of Beringia. Chicago: University of Chicago Press, xxi, 576 p.
Notas
Quienes defienden esta hipótesis ven cualquier afirmación de una colonización de América antes de esa fecha con mucho escepticismo. ↩
Aunque, por el contrario, se argumenta que ese vacío de 16000 años en el registro arqueológico se debe a que los yacimientos que podrían arrojar luz sobre esta cuestión están bajo el agua. ↩
La política, la geografía y la tradición han centrado durante mucho tiempo la atención arqueológica en la evolución de Homo sapiens en Europa y África. Ahora, una nueva investigación desafía las viejas ideas al mostrar que las primeras migraciones humanas se desarrollaron en Asia mucho antes de lo que se sabía con anterioridad.
El desierto de Nefud es una zona desolada de dunas de arena anaranjadas y amarillas. Cubre aproximadamente 40000 km cuadrados de la Península Arábiga. Pero hace decenas de miles de años, esta área era una tierra exuberante de lagos, con un clima que podría haber sido más agradable para la vida humana.
Una tarde de enero de 2016, un equipo internacional de arqueólogos y paleontólogos estudiaba la superficie del antiguo lecho de un lago en un yacimiento llamado Al Wusta en el paisaje de arena y grava del Nefud. Buscaban atentamente fósiles, trozos de herramientas de piedra y cualquier otro signo que hubiera quedado del pasado alguna vez verde de la región.
De repente, Iyad Zalmout, un paleontólogo que trabaja para el Servicio Geológico de Arabia Saudi, localizó lo que parecía un hueso. Con pequeños picos y pinceles, él y sus colegas extrajeron el hallazgo del suelo.
«Sabíamos que [era] importante», recordó Zalmout en un correo electrónico. Era la primera prueba directa de la presencia de primates u homínidos en el área. En 2018, las pruebas de laboratorio confirmaron que este espécimen era el hueso de un dedo de un humano anatómicamente moderno que habría vivido hace al menos 86000 años.
Antes de este descubrimiento de Al Wusta, las pruebas en forma de herramientas de piedra habían sugerido la presencia humana en el Nefud hace entre 125000 y 55000 años. Para los antropólogos, «humano» y «hominino» pueden significar cualquiera de varias especies estrechamente relacionadas con la nuestra. El hueso del dedo era el Homo sapiens más antiguo encontrado en la región.
La datación del hueso contradice un discurso bien establecido en la comunidad científica. Los hallazgos, particularmente en el área de los actuales Israel, Jordania y Líbano –conocidos como el «Levante»-, han llevado a comprender que H. sapiens salió por primera vez de África no antes de hace 120000 años, probablemente migrando hacia el norte junto a la costa mediterránea. Estas personas se establecieron en el Levante y sus descendientes –o aquellos de una posterior migración humana fuera de África–, viajaron a Europa decenas de miles de años después.
Solo más tarde, sigue la historia, viajaron a partes de Asia, como Arabia Saudí. Según algunas estimaciones, los humanos anatómicamente modernos no habrían estado en lo que ahora es Al Wusta hasta hace unos 50000 años.
El hueso del dedo, por lo tanto, supone un nuevo giro en la historia de cómo y cuándo nuestra especie dejó el continente africano y, a trompicones, pobló gran parte del resto de la tierra. Una nueva serie de descubrimientos, particularmente en Asia, sugieren que los humanos modernos abandonaron por primera vez África hace unos 200000 años, tomando muchas rutas diferentes.
El Levante ya no es necesariamente central, y los lugares al este podrían haber tenido una importancia inesperada en las primeras migraciones humanas. Como dice el antropólogo Michael Petraglia, del Instituto Max Planck para la Ciencia de la Historia Humana, «se está revelando una nueva historia».
Estos hallazgos podrían arrojar luz sobre grandes preguntas sin respuesta, como por qué los humanos hicieron estas migraciones, cuáles fueron las condiciones ambientales del pasado, y cómo interactuó H. sapiens con otros homininos. Pero esta nueva narrativa también subraya cuánto de nuestro conocimiento proviene, y está limitado por, dónde han trabajado los arqueólogos y otros investigadores. El énfasis geográfico ha estado influenciado durante mucho tiempo no por la ciencia sino por el acceso, la financiación y la tradición.
El primer indicio de que la larga historia de los viajes humanos fuera de África había pasado por alto algo crítico provino de la región del Levante, concretamente de la Cueva Misliya en Israel. En 2018, los arqueólogos anunciaron que habían encontrado una mandíbula humana en esta cueva.
El hueso –datado con tres métodos diferentes en el curso de una investigación que ha durado diez años–, tiene entre 177000 y 194000 años, adelantando la cronología de la presencia humana allí al menos 50000 años. Y unas herramientas de piedra más antiguas halladas bajo la mandíbula sugieren que los humanos podrían haber estado en esa área incluso antes.
Entonces, es posible que los humanos abandonaran África y viajaran al Levante, y a otros lugares, incluso antes de la fecha que indica esa mandíbula. Esta idea ganó aún más fuerza en julio de 2019, cuando un grupo de investigadores publicó nuevos hallazgos acerca de un cráneo descubierto en Grecia en la década de 1970. Ese fósil, sugiere el nuevo estudio, es humano y tiene más de 210000 años.
Pero además de esta cronología cambiante, los investigadores están replanteándose a dónde viajaron los humanos cuando salieron de África. El hallazgo de Al Wusta es solo un ejemplo.
En 2015, investigadores de China publicaron el hallazgo de 47 dientes humanos, con una antigüedad de entre 85000 y 120000 años, en una cueva de la provincia de Hunan. Hasta este descubrimiento, los fósiles de seres humanos modernos más antiguos encontrados en el sur de Asia tenían solo unos 45000 años.
Estos nuevos hallazgos «obligan a replantearnos cuándo y cómo nos dispersamos», dice la antropóloga forense María Martinón-Torres, directora del Centro Nacional de Investigación sobre la Evolución Humana en Burgos, España, y miembro del equipo que descubrió y estudió los dientes. Añade: «Pudo haber más de una migración “fuera de África”… los humanos, como cualquier otro animal, pudieron haberse expandido hasta que no hubo ninguna barrera, ecológica o geográfica, que les impidiera hacerlo».
En 2018, investigadores de la India publicaron el descubrimiento de una colección de herramientas de piedra avanzadas. Afirman que este hallazgo indica la presencia de homininos que se remontaría al menos 170000 años, miles de años antes de lo sugerido por las investigaciones anteriores. Y otras pruebas sugieren que los primeros humanos pudieron haberse dirigido directamente a Asia al cruzar desde África a lo largo de la Península Arábiga, rodeando el Levante, de donde proviene gran parte de las pruebas más antiguas de los seres humanos fuera de África.
Una serie de nuevos descubrimientos ha cambiado la comprensión del tiempo, las rutas y el alcance geográfico asociado con la dispersión de H. sapiens fuera de África. Pero para los arqueólogos, estos hallazgos también señalan una especie de punto ciego. Como dice Martinón-Torres, «estos hallazgos también son una llamada de atención con respecto a Asia».
De hecho, existe una mayor conciencia acerca de la necesidad de ampliar el ámbito geográfico de la paleontología y la arqueología relacionadas con las primeras migraciones humanas y la evolución. «Durante mucho tiempo», agrega Martinón-Torres, «Asia se consideró como un callejón sin salida con un papel secundario en la corriente principal de la evolución humana».
«Hay un enorme sesgo en el trabajo de campo arqueológico y dónde se está produciendo, y nuestras teorías sobre la evolución humana se basan en estos sesgos geográficos», dice Petraglia, quien junto con Zalmout y sus colegas de la Comisión Saudita de Turismo y Patrimonio Nacional encontraron el hueso de Al Wusta.
Varios factores han contribuido a este sesgo, explica la arqueóloga y escritora Nadia Durrani, coautora del libro «Archaeology: A Brief Introduction» junto al antropólogo Brian Fagan. La arqueología comenzó hace más de un siglo «como una disciplina científica occidental», dice.
Los primeros arqueólogos, que eran europeos y estadounidenses, se centraron principalmente en la Europa mediterránea y los lugares mencionados en la Biblia, incluidos los actuales Irán, Irak, Egipto, Israel y Cisjordania. «La gente estaba interesada en la Biblia y los temas clásicos», incluida la antigua Grecia y Roma, dice Durrani. A medida que los arqueólogos hicieron descubrimientos en esas áreas, el interés en esas regiones creció y las instituciones también se asentaron en esos lugares, lo que a su vez alimentó más investigaciones allí.
«Los países donde se ha venido desarrollando durante décadas la investigación paleoantropológica tienen más probabilidades de tener hallazgos importantes, que son tanto bien conocidos como valorados», dice Katerina Harvati, directora de paleoantropología de la Universidad de Tübingen. «Y por lo tanto, es probable que tengan más oportunidades de financiación».
Lo contrario también es cierto. Puede ser difícil convencer a colegas o a posibles financiadores del potencial de un lugar cuando se ha explorado poco y carece de ciertas infraestructuras. Las barreras ambientales y naturales también juegan su papel. Petraglia señala que trabajar en áreas que no han sido bien exploradas puede exigir comenzar desde el principio, con tareas como reconocimientos y mapeo, y a menudo no hay un trabajo previo en el que basarse.
Por ese motivo, los problemas políticos pueden ayudar u obstaculizar a los arqueólogos. Durrani, por ejemplo, llevó a cabo trabajo de campo en Yemen en la década de 1990, y luego dirigió visitas guiadas a yacimientos arqueológicos. Tuvo que detener su trabajo en 2008 debido a la inestabilidad política en la zona. La violencia y los conflictos suponen graves obstáculos de acceso, dice.
Con el tiempo, los científicos esperan que el acceso y las condiciones mejoren aún más. Mientras tanto, esta investigación muestra que los humanos anatómicamente modernos abandonaron África antes de lo esperado y viajaron hacia el sur, a lo largo de la Península Arábiga, además de hacia el norte.
Sin embargo, algunos de estos hallazgos han generado escepticismo. Jeffrey Schwartz, profesor emérito de la Universidad de Pittsburgh, advierte en contra de extraer conclusiones dramáticas de los hallazgos. «Creo que estamos llamando a demasiadas cosas H. sapiens», dice.
Por el contrario, Mina Weinstein-Evron, una arqueóloga de la Universidad de Haifa que fue codescubridora de la mandíbula de la Cueva de Misliya, sospecha que los hallazgos recientes pertenecen a H. sapiens, aunque está de acuerdo en que la historia de la migración de los humanos anatómicamente modernos dista de ser clara. «No sabemos nada. Tenemos una prueba aquí y otra allá», dice. «Y luego usamos grandes palabras como “migración” y “dispersión”. Hablamos como si hubieran comprado un billete. Pero no sabían a dónde iban. Para ellos quizás ni siquiera fuera un movimiento, tal vez viajaron 10 kilómetros cada generación».
Además, algunos hallazgos genéticos sugieren que, incluso si los humanos viajaron fuera de África hacia Asia antes de lo que se pensaba anteriormente, es posible que esas primeras migraciones no hubieran tenido éxito desde una perspectiva evolutiva. Según las conclusiones de tres grupos diferentes de científicos publicadas en Nature en 2016, el ADN de los euroasiáticos divergió del de los africanos hace entre 60000 y 80000 años. En otras palabras, todos los humanos vivos hoy en día son descendientes de H. sapiens que emigraron fuera de África dentro de esa ventana, así como otros homininos, como los neandertales.
Posibles rutas de migración de Homo sapiens. Tomado de: Bae, C. J.; Douka, K. y Petraglia, M. D. (2017), «On the origin of modern humans: Asian perspectives». Science, vol. 358, núm. 6368, p. eaai9067.
No obstante, las migraciones anteriores son intrigantes, dice Luca Pagani, un antropólogo biológico que escribió uno de los artículos de Nature. «Aunque no va a cambiar nuestra idea de cuáles migraciones fueron exitosas, muestra una variedad más rica de intentos de dispersión», dice, y esa es una parte esencial de la historia de los primeros humanos modernos.
De hecho, las razones por las que fallaron algunas migraciones de los primeros humanos podrían aclarar algunas de las principales cuestiones en arqueología. Martinón-Torres y sus colegas que trabajan en China, por ejemplo, han postulado que los primeros humanos modernos podrían haber competido con los neandertales u otros homininos, lo que podría haber influido en sus desplazamientos.
Petraglia, por otra parte, sospecha que los primeros humanos modernos pueden haber prosperado en el yacimiento árabe hasta que el agua desapareció a medida que el desierto crecía. «Si quieres saber cómo puede afectarnos el cambio climático algún día, bueno, aquí tenemos una historia completa sobre los efectos del cambio climático en las poblaciones humanas», dice. En resumen, puede que los descendientes de estos intrépidos humanos no hubieran sobrevivido, pero sus historias aún podrían guiarnos hacia el futuro.
Este trabajo apareció publicado por primera vez en SAPIENS bajo una licencia CC BY-ND 4.0. Lee el original aquí.
[…] así como rechazamos una medicina amarga, nos resistimos a aceptar que el sufrimiento es esencial a la vida, de modo que no fluye hacia nosotros desde fuera, sino que cada uno lleva la fuente inagotable del mismo en su propio interior.
En la Grecia clásica, con Platón, Demócrito y Alcmeón de Crotona, se defendía que el cerebro debía considerarse el centro de la razón y de las sensaciones, incluida la dolorosa. De hecho, Aristóteles definió las emociones básicamente como afecciones acompañadas de placer o dolor 2. Descartes, por su parte, contemplaba la experiencia dolorosa a partir del concepto dual de la naturaleza humana (que distinguía entre el cuerpo y el alma). Para él el dolor era una percepción del alma, que podía estar originada por la acción de agentes externos al cuerpo, o bien por el propio cuerpo. De esta forma, la biología tenía la capacidad de estudiar el «mecanismo» del dolor, describir objetivamente su origen, el recorrido y el destino del estímulo doloroso.
Solemos experimentar el dolor de forma transitoria cuando lo provoca alguna enfermedad o un traumatismo. Sin embargo, para muchas personas el dolor es crónico, se ha convertido en un «compañero» que no desaparece ni con los analgésicos 3 más potentes. La situación no es sencilla ya que el dolor es un complejo «sistema de defensa», que nos avisa de que algo no funciona, y cuya percepción es subjetiva, es desagradable y posee una importante carga emocional.
La Asociación internacional para el estudio del dolor (IASP, por «International Association for the Study of Pain») puntualiza que existen dos posibles fuentes del dolor: una emocional y otra sensorial. Así, la IASP define el dolor como «una experiencia sensorial y emocional desagradable asociada con un daño tisular real o potencial, o descrita en términos de dicho daño 4. A partir de esta definición, podemos analizar sus dos componentes principales:
El componente sensorial. Proporciona la información nociceptiva como consecuencia de la transmisión del estímulo lesivo al córtex cerebral (se trata de la sensación dolorosa en sí). Distinguimos entre el «dolor nociceptivo», que es el causado por la activación de los receptores del dolor (son los «nociceptores», de los que hablaremos más adelante) en respuesta a un estímulo (lesión, inflamación, infección, enfermedad); mientras que el «dolor neuropático» se origina por un estímulo directo del sistema nervioso central o una lesión de los nervios periféricos.
El componente emocional. Confiere al dolor un carácter único y personal –subjetivo– al originar una respuesta emocional y de conducta secundaria a la sensación dolorosa. Aquí debemos mencionar el «dolor idiopático», en el que predominan los factores psicosociales, y que suele ser característico de dolores crónicos.
Para completar lo dicho hasta ahora, me gustaría añadir otra definición –quizás más amplia– del término «dolor» que encontramos en el diccionario Dorland de medicina, y que recoge dos acepciones: 1. Uno de los signos cardinales de la inflamación; y 2. Sensación más o menos localizada de malestar, molestia o aflicción, que resulta de la estimulación de terminaciones nerviosas especializadas. Actúa como mecanismo de protección, en la medida en que induce a quien lo sufre a retirarse o apartarse de la fuente.
Clasificación del dolor y analgésicos recomendados. Fuente: Kim, et al. (2020), «All about pain pharmacology: what pain physicians should know».
La mayoría de textos que tratan sobre el dolor lo describen como un «síntoma». De hecho, la literatura recoge que se trata del principal motivo que lleva a los pacientes a acudir a los centros de atención sanitaria.
A pesar de que contamos con una variada gama de medicamentos para su tratamiento, lo cierto es que en las dos últimas décadas no se ha desarrollado un solo analgésico realmente innovador [ 1. Ver en este sentido Correll y Kissin. (2019).]. Muchos pacientes sufren esa carencia, mientras que otros deben hacer frente a los importantes efectos secundarios de aquellos con los que contamos y que complican los tratamientos prolongados. Por todo ello, el estudio de la biología del dolor y la búsqueda de medios más eficaces para el tratamiento del dolor crónico son ahora más urgentes que nunca.
Los trabajos de los científicos se están centrando, fundamentalmente, en dos parcelas relacionadas:
En primer lugar, existe un interés por comprender mejor el mecanismo por el que las señales dolorosas viajan desde los nervios hasta el cerebro, y la forma en que éste percibe esa sensación de dolor.
En segundo lugar, aumentan los estudios que tratan de desvelar el papel que algunos genes desempeñan en la regulación del dolor. Siguiendo esta vía se persigue explicar por qué algunas personas sienten y toleran el dolor de forma tan diferente.
Sintiendo el dolor
[…] limitémonos, pues, a hablar del dolor. Concedo de buen grado que sea la desgracia mayor de nuestra vida, pues soy de los que más la detestan y de los que más le huyen, por no haber tenido hasta el presente, gracias Dios, gran comercio con él; yo creo que en nosotros reside, si no el poder de reducirlo a la nada, al menos el de debilitarlo por la paciencia, y el de alcanzar, a pesar de los sufrimientos corporales, que el alma y la razón se mantengan resistentes y bien templadas.
«Ensayos», Michel de Montaigne, Libro I, capítulo XL.
Como hemos indicado, la capacidadde sentir dolor es un mecanismo de defensa del organismo que actúa para disminuir el riesgo de sufrir lesiones. Se trata de un sistema de alarma clave para la autopreservación ya que gracias a él somos conscientes de los peligros que amenazan nuestra integridad física. Para comprender su importancia, basta con imaginarnos lo complicada que sería nuestra vida sin él. Por ejemplo, la «insensibilidad congénita al dolor» es un conjunto de enfermedades hereditarias poco frecuentes que se caracterizan porque las personas afectadas no sienten el dolor. Los enfermos sufren quemaduras importantes (dado que no se percatan del daño en su piel), lesiones graves de huesos por fracturas que no detectan, úlceras en la piel por el roce o contacto continuo con superficies abrasivas, etc.
Sin embargo, el dolor no sólo puede ser «sensorial», también hay que tener presente el dolor «emocional». Este tipo de dolor se puede manifestar de muchas maneras y precisa de un abordaje especial. Por ejemplo, el compositor finlandés Jean Sibelius escribió en 1900 su Opus 20 para violonchelo y piano, titulada Malinconia, tras el fallecimiento de Kirsti, su hija más pequeña 5, a consecuencia de una epidemia de tifus. Oyendo esta pieza podemos «sentir» realmente el dolor del compositor en los «largos, desencajados e insistentes pasajes de piano y el tono intensamente sombrío e incluso irritante del violonchelo». La melancolía que emana de esta pieza sin duda nos perturba y provoca un peculiar cúmulo de emociones.
Ahora bien, siendo evidente que el dolor tiene una parte positiva, también es cierto que muchas personas sufren cuando el dolor provocado por una lesión se prolonga mucho tiempo después de que se haya resuelto la causa que lo ha originado. En estos casos el dolor se convierte en una enfermedad en sí misma.
Veamos por tanto con más de detalle cómo «sentimos» el dolor.
Acciones eferentes de los nociceptores. Un estímulo nocivo conduce a potenciales de acción en fibras nociceptivas que se propagan no sólo al sistema nervioso central sino también antidrómicamente hacia las ramas periféricas. Estos potenciales de acción antidrómica producen la liberación de neuropéptidos, como sustancia P, péptido relacionado con el gen de calcitonina (CGRP) y neurocinina A (NKA). Estas sustancias pueden estimular a las células epidérmicas (1) y a las células inmunes (2) o producir vasodilatación (3), extravasación plasmática (4) y contracción del músculo liso (5). Tomado de McMahon et al. (2007).
Las señales dolorosas viajan desde la piel o los órganos internos hasta la médula espinal a través de una clase especial de neuronas sensitivas llamadas «nociceptores». Sus fibras se extienden hasta la piel, los pulmones, los intestinos y otras partes del cuerpo, y están equipadas para percibir distintos tipos de estímulos nocivos. En este sentido, la naturaleza ha adoptado una estrategia mixta. Algunos nociceptores responden a diferentes tipos de estímulos térmicos y mecánicos, y transmiten con rapidez la señal a través de fibras mielinizadas; otros en cambio están conectados a las fibras amielínicas –de transmisión más lenta– que responden a la presión, temperatura y otro tipo de estímulos (así se explican las diferentes sensaciones que producen una quemadura, una punción o un pinchazo).
Cuando estas neuronas detectan una amenaza (un corte, una quemadura etc.) envían señales eléctricas a la médula espinal. Son las fibras aferentes las que penetran en la médula espinal a través de las raíces dorsales, y terminan en las astas posteriores de la sustancia gris. Ahí contactan con las neuronas medulares que llevan la señal a las regiones cerebrales donde se transforma esa información en la percepción del dolor 6.
El cerebro, al registrar el dolor, reacciona con una señal analgésica para calmarlo. Se trata de la «vía descendente del dolor» por la que las redes neuronales del cerebro envían señales eléctricas a través de la médula espinal que desencadenan la segregación de endorfinas y otros opioides naturales. Estas sustancias bioquímicas inhiben las señales ascendentes con lo que reducen el grado de dolor percibido.
El descubrimiento de la «sensibilización central» 7, un fenómeno según el cual podría existir dolor en ausencia de una lesión desencadenante, echó por tierra la opinión de algunos médicos de que los pacientes que se quejaban de dolor sin que mediase una lesión obvia debían estar mintiendo.
Esto sucede porque el sistema de transmisión del dolor puede hipersensibilizarse a raíz de una lesión; aunque también puede descontrolarse sin razón o quedarse atrapado en un estado hipersensible mucho después de la curación de la lesión original, con lo que la señal analgésica enviada por el cerebro no consigue contrarrestar el dolor. Hablamos del dolor neuropático, la fibromialgia etc. Como hemos comentado, para estos pacientes el dolor no es un síntoma, sino una enfermedad causada por el mal funcionamiento del sistema nervioso.
Tratando el dolor
No existe objetivo más noble que aliviar el dolor y el sufrimiento.
«Tratado del dolor». Ronal Melzack.
Llegados a este punto vamos a analizar algunos de los pasos que se están dando para mejorar el abordaje terapéutico del dolor.
Diseño de fármacos que impidan la transmisión de la señal dolorosa.
Varios investigadores trabajan en el diseño de un medicamento que impida o module la transmisión de la señal del dolor. Bastaría, por ejemplo, con bloquear los receptores de glutamato, el neurotransmisor que utilizan las neuronas nociceptivas. Este neurotransmisor juega un papel importante en la modulación del dolor ya que es el principal aminoácido excitatorio del sistema nervioso central (en su condición de principal responsable de la transmisión sináptica rápida). Y en este sentido estamos de suerte ya que contamos con un fármaco que hace precisamente eso: la ketamina.
El efecto analgésico de la ketamina se conoce desde su introducción en 1966 como un anestésico general. Sin embargo, ahora se ha descubierto un nuevo objetivo molecular, concretamente su función como un antagonista del receptor NMDA (receptor excitatorio del Glutamato N-metil D-aspartato). Pese a que la ketamina se emplea cada vez más como un analgésico para el tratamiento del dolor agudo y crónico, presenta un grave inconveniente: a dosis efectivas posee efectos secundarios muy severos (entre los que se encuentran sedación, náuseas, reacciones disociativas, modificación de la conducta etc.).
Por esa razón se busca otro enfoque. Un grupo de investigadores liderado por Lucía Hipólito sabía que esos receptores NMDA se encuentran regulados a su vez por otras proteínas de la membrana de la neurona, los llamados canales pequeños de potasio (SK channels). Lo que este grupo ha logrado –en modelos de ratón– ha sido obtener un efecto analgésico potente sin los efectos secundarios ya conocidos, gracias a la combinación de fármacos que interaccionan con los canales SK a dosis subefectivas (es decir, en dosis que no provocan ningún efecto observable), con dosis subefectivas de fármacos tipo ketamina 8.
A pesar de lo prometedor de este trabajo, problemas de financiación han impedido a la Dra. Hipótito continuar con esta investigación. Esperemos que la situación cambie, o que otro grupo tome el relevo y siga los pasos de esta investigadora.
Empleo del estado emocional de los pacientes para controlar el dolor.
Como hemos visto a lo largo de esta anotación, las investigaciones sobre el dolor han confirmado que se trata de un fenómeno complejo y subjetivo, y que la forma en que sentimos el dolor depende tanto del cerebro de la persona que lo experimenta, como del momento en que lo hace. Esto significa que la traducción de las señales de los nociceptores en sensaciones dolorosas se ve influida por el estado emocional de la persona. Al mismo tiempo, el contexto emocional en el que se percibe el dolor también puede alterar cómo se experimenta (un ejemplo de esta situación sería el dolor o las molestias que tienes después de una sesión de ejercicio intenso. Es una sensación dolorosa, pero placentera al mismo tiempo).
Ahora sabemos que en el cerebro no existe un centro del dolor como tal, sino que diferentes regiones se activan como reacción a los estímulos dolorosos. Entre ellas se incluyen las redes que también participan en la emoción, la cognición, la memoria y la toma de decisiones. Es decir, un mismo estímulo puede generar un patrón de activación distinto según el momento en que se produzca, de ahí que la experiencia personal del dolor pueda variar aunque el daño sea parecido.
La neurocientífica de la Universidad de Oxford Irene Tracey y su equipo, han planteado un enfoque innovador para sacar partido a esta situación. Diseñaron un experimento para el que reclutaron a un grupo de pacientes con dolor neuropático tras una toracotomía. A todos se les dijo que recibirían un nuevo tratamiento para el dolor, cuando lo cierto es que la mitad de ellos no recibieron esa medicación. El alivio que experimentaron los pacientes de ese segundo grupo no fue más que el resultado del efecto placebo.
Es decir, estamos ante un caso en el que la mera expectativa de ver reducido el dolor se traduce en un alivio auténtico. Parece ser que el placebo activa la «vía descendente», que provoca la liberación de opioides sintetizados en el cerebro.
Otro enfoque para el control emocional del dolor es el seguido por el equipo del Dr. Andre Machado9. El Dr. Machado y su equipo implantaron microelectrodos en una zona del cerebro relacionada con el procesamiento de las emociones.
Se trata de un procedimiento similar a la estimulación cerebral profunda, utilizada para tratar algunos síntomas de la enfermedad de Parkinson como el temblor, rigidez, movimiento lento etc. Consiste en la estimulación eléctrica leve de alta frecuencia en una zona específica del tálamo, el globo pálido o el núcleo subtalámico por medio de un electrodo implantado en el encéfalo. Su función es enviar una señal eléctrica a estas áreas específicas del cerebro que controlan el movimiento, bloqueando de esta forma las señales nerviosas anormales.
Los hallazgos sugieren que el cerebro de los pacientes con dolor crónico está condicionado por la exposición constante al dolor y reacciona como si todos los estímulos fuesen potencialmente dolorosos. Este tipo de tratamiento, que aún no está autorizado de forma generalizada, permite que el cerebro empiece a distinguir de nuevo lo doloroso de lo que no lo es.
La genética también juega su papel.
Por último, los estudios genéticos también están aportando su grano de arena para comprender tanto los procesos dolorosos, como potenciales vías terapéuticas.
James Cox, genetista del University College, ha descubierto que los genes FAAH y FAAH-OUT, presentan mutaciones que reducen la descomposición de un neurotransmisor llamado anandamida, lo que contribuye a aliviar el dolor 10. Es decir, aquellas personas con esas mutaciones poseen un exceso de esta sustancia bioquímica, lo que las vuelve prácticamente inmunes al dolor (algo que hemos visto que no es tan positivo como podría llegar a pensarse). Actualmente hay varios estudios en marcha que tratan de poner a prueba diversas posibilidades para el uso terapéutico de este hallazgo.
Otro gen, SCN9A, también está implicado en la señalización del dolor. SCN9A codifica para una proteína que resulta clave en la transmisión de las señales desde las neuronas nociceptivas hasta la médula espinal. Una proteína, Nav1.7, se sitúa sobre la superficie de la neurona y hace las veces de canal para que los iones de sodio accedan al interior, permitiendo que los impulsos eléctricos de la señal dolorosa se transmitan. Las mutaciones en este gen producen versiones deformes de esta proteína, que no pueden cumplir su función. Si por ese motivo los nociceptores no pueden de conducir las señales de dolor, los pacientes no lo sienten.
Dado que esta proteína Nav1.7 sólo está presente en las neuronas sensibles al dolor, si podemos desarrollar un medicamento que cierre selectivamente ese canal, podría convertirse en un analgésico eficaz. Hasta el momento, el único efecto secundario conocido de esta condición es la pérdida de olfato.
Queda aún mucho camino para lograr verdaderos avances en la clínica, es decir, en el tratamiento eficaz y sin efectos secundarios del dolor de tantos y tantos pacientes que sufren a diario. Sin embargo, tampoco podemos dejar de lado que hay un buen número de investigadores en todo el mundo dedicados a esta tarea.
Beard, D. J., et al. (2020), «Considerations and methods for placebo controls in surgical trials (ASPIRE guidelines)». The Lancet, vol. 395, núm. 10226, p. 828-838.
Correll, D. J. y Kissin, I. (2019), «Problems with developments of breakthrough analgesics: recent history via scientometric analysis». Journal of Anesthesia History, vol. 5, núm. 2, p. 49-57.
Dorland, W. A. N. (2005), Dorland diccionario enciclopédico ilustrado de medicina. Madrid [etc.]: McGraw-Hill Interamericana de España, xxvii, 2210 p.
Fue el poeta y médico norteamericano Oliver Wendell Holmes quien sugirió que a los compuestos que suprimían el dolor se los denominara «anestésicos» (de las palabras griegas que significan «sin sensación»). Por su parte, la «analgesia» (una palabra que también proviene del griego) se define por la RAE como la «falta o disminución de las sensaciones dolorosas, que no afecta a los demás sentidos». ↩
Traducción propia de: «An unpleasant sensory and emotional experience associated with actual or potential tissue damage, or described in terms of such damage». Ver la terminología adoptada por la IASP aquí. En una nota a dicha definición se indica que el hecho de que dolor crónico se considere como una enfermedad o un trastorno en sí mismo no hace necesario un cambio en la definición. ↩
Falleció el 13 de febrero con solo quince meses de edad. ↩
Además de estos, existen otros circuitos neuronales que intervienen en la modulación del dolor y que están implicados en sus factores afectivos y emocionales. ↩
Cuando un tejido se daña y el dolor persiste durante unos días, se producen unos mecanismos de adaptación de los nociceptores y de la respuesta nociceptiva al daño. Este proceso se llama «hiperalgesia primaria» o sensibilización periférica, y representa una acción protectora del cuerpo humano frente a posibles daños futuros. La «hiperalgesia secundaria» se refiere al proceso de incremento de la respuesta en el asta dorsal, localizado en los segmentos del procesamiento de la nocicepción. Mientras que la sensibilización periférica es temporal, la hiperalgesia secundaria o «sensibilización central» es un proceso que engloba al sistema nervioso central. ↩
Nombre taxonómico: Homo luzonensis ―Détroit, F., et al. (2019)―. Descubiertos en la cueva de Callao —en la isla de Luzón— el 24 de agosto de 2011, los fósiles pertenecen al menos a dos adultos y un niño. Antigüedad: entre 67000 y 50000 años.
Espécimen tipo: CCH6 (conjunto de cinco fósiles: CCH6-a a CCH6-e), que se corresponde a la dentición postcanina del maxilar de un único individuo.
Características y comportamiento: Los fósiles de Luzón presentan un conjunto único de rasgos físicos que los diferencia del resto de congéneres que vivían en esa misma época. Algunas de estas características parecen muy primitivas —como, por ejemplo, el pequeño tamaño y la forma sencilla de las coronas de los molares; o la curvatura de los dedos de las manos y los pies, que lo acercan más a los australopitecinos—; mientras que otras —en especial sus dientes— son similares a los parántropos, Homo erectus e incluso Homo sapiens.
Puesto que sus manos y pies son más primitivos que los de Homo erectus se plantea la cuestión de si el antepasado de Homo luzonensis sería incluso más antiguo que Homo erectus y si, por tanto, salió fuera de África antes de que lo hiciera aquél. Dada la antigüedad y localización de los fósiles, también surge la cuestión de la forma en que pudo llegar a Filipinas.
La pregunta de si algún hominino había salido de África antes de que lo hiciera Homo erectus ya se planteó cuando se descubrió Homo floresiensis. Ahora es una hipótesis que cobra más fuerza.
La cueva de Callao, en la isla filipina de Luzón, supone la más importante atracción turística de una región horadada por más de 300 cuevas. Además de constituir una fuente de ingresos, también se llevan a cabo excavaciones arqueológicas de forma ininterrumpida desde 2007, año en el que se descubrió una falange catalogada como CCH1 1, por Callao Cave Hominin 1, o «hominino 1 de la cueva de Callao».
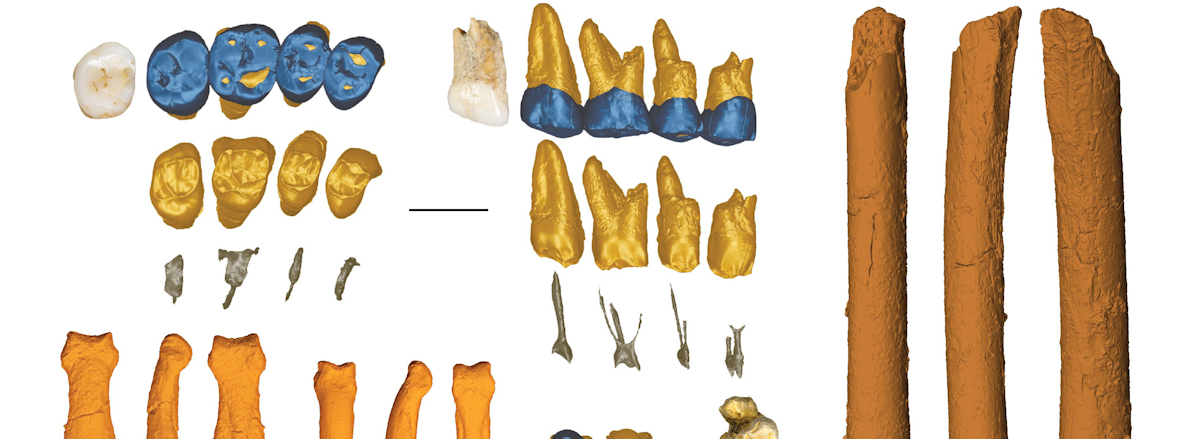
Sucesivas campañas han permitido recuperar un total de 12 nuevos restos 2 que presentan una mezcla única de caracteres primitivos (es decir, rasgos más parecidos a los australopitecinos) y derivados (similares a los de Homo sapiens por ejemplo) que hace que sus descubridores los asignen a un nuevo miembro del género Homo: Homo luzonensis.
En un artículo publicado en la revista Nature el 10 de abril de 2019 los investigadores describen estos restos y confirman que pertenecieron a tres individuos: dos adultos y un niño 3. La datación del fósil más antiguo (CCH1), mediante la técnica de series de Uranio arroja una antigüedad mínima de 67000 años.
Caracteres
Dado que no contamos con fragmentos del cráneo no podemos hacer suposiciones acerca de su aspecto; y dado que el fémur recuperado está fracturado, tampoco podemos saber su altura.
Los dientes son «sencillos», con pocas protuberancias. Comparados con la dentición de Australopithecus, Paranthropus y otras especies del género Homo, los dientes de H. luzonensis son pequeños y están comprimidos mesiodistalmente. Además, hay un marcado contraste entre su tamaño relativo y la forma de los premolares y molares. En este sentido, los premolares de H. luzonensis tienen un tamaño relativo en relación a los molares muy grande, más que en el caso de H. floresiensis. Tienen varias raíces (P3 tiene tres, P4 tiene dos) muy robustas y divergentes, una característica arcaica que es típica de Australopithecus y Paranthropus. Así, los premolares combinan unos caracteres de tamaño y forma vistos en especies como H. neanderthalensis, H. floresiensis y H. sapiens. Por otro lado, los molares son extremadamente pequeños y se parecen mucho a los de H. sapiens y, en menor medida, a los de H. floresiensis.
En definitiva, el patrón de la morfología de los dientes de Homo luzonensis es fácilmente distinto, y distinguible, del resto de especies analizadas.
Conjunto dientes CCH6.
Por otro lado, los huesos de las manos y los pies son mucho más primitivos, comparables a los de los australopitecos que vivían en África alrededor de dos millones de años antes, y cuyas extremidades estaban adaptadas para subir a los árboles.
Por ejemplo, CCH2, una falange intermedia de la mano, es larga, estrecha y con una curva muy marcada. Presenta por tanto unos caracteres primitivos similares a los observados en Australopithecus y, en menor medida, en el holotipo de Homo habilis (OH 7), pero no en otras especies posteriores del género Homo (aunque en algunos aspectos sea similar a las falanges de Homo naledi). Por su parte, CCH4, una falange del pie, presenta unos caracteres primitivos similares a los de Australopithecus (en una combinación de varias especies), y concretamente es indistinguible de las vistas en Au. afarensis y Au. africanus.
CCH2. Falange intermedia de la mano.
Por el momento, probablemente sea razonable aceptar la nueva especie, al menos provisionalmente, mientras se esperan más hallazgos.
¿Qué implicaciones tiene Homo luzonensis para la evolución humana?
Los autores admiten que establecer el origen de Homo luzonensis, así como sus relaciones filogenéticas con otros homininos, son cuestiones que quedan sin respuesta por el momento. Secuenciar su ADN ayudaría a clarificar el panorama, pero los intentos de extraer el material genético de los restos recuperados han resultado infructuosos debido a la humedad y la elevada temperatura de las islas del Sudeste Asiático.
A pesar de todo, con los datos de que disponemos es posible aventurar algunas hipótesis.
Sabemos que la isla de Luzón ha estado rodeada por mar desde hace dos millones y medio de años. Por lo tanto, es evidente que H. luzonensis ha tenido que llegar allí usando algún tipo de embarcación. De nuevo nos topamos con el mismo interrogante que se planteó cuando se encontraron los restos de H. floresiensis y que generó —y sigue generando— intensos debates.
También debemos señalar que en el yacimiento arqueológico de Kalinga, en un valle cercano a la cueva de Callao, se han encontrado herramientas de piedra y huesos de un rinoceronte con marcas de corte en las costillas y en las extremidades, así como de percusión en los huesos de una de las extremidades anteriores 4. Este hallazgo, con una antigüedad de más de 700000 años, confirma la presencia de homininos en la región en una época tan temprana. Aunque es posible que quien se alimentara del rinoceronte fuera H. luzonensis, aún no contamos con pruebas contundentes al respecto.
Llegados a este punto, los especialistas plantean al menos cuatro diferentes posibilidades acerca de su origen:
Por lo que sabemos a día de hoy, sólo dos especies se aventuraron fuera de África hasta llegar al Sudeste Asiático: H. erectus, hace entre 2 y 1,5 Ma; y H. sapiens, que hace alrededor de 60000 años ya había llegado hasta Australia. Una posibilidad por tanto es que H. luzonensis fuera descendiente de los primeros H. sapiens que salieron de África y que sabemos con seguridad que poseían los conocimientos y la capacidad técnica para navegar por el océano. Tras llegar a Luzón, quizá evolucionaron de forma acelerada al carecer de intercambio genético con otros humanos, lo que hizo que presenten la extraña mezcla de rasgos que vemos ahora.
Otra posibilidad es que descienda de los H. erectus que de algún modo llegaron a Luzón en su camino a lo que hoy es China. Al igual que H. floresiensis, representarían los últimos supervivientes de esa migración, y habría evolucionado durante decenas de miles de años aislado con las presiones evolutivas que eso supone.
La tercera opción es que H. luzonensis provenga de una oleada que salió de África antes que H. erectus, posiblemente de algún australopitecino. Ahora mismo no hay fósiles ni datos que permitan sostener esta hipótesis.
La cuarta opción, defendida por Chris Stringer, es que tanto H. luzonensis como H. floresiensis descienden de un antepasado común que pudo surgir en la isla de Sulawesi por ejemplo, donde se han hallado herramientas de piedra de unos 110000 años de antigüedad 5.
Las excavaciones en la cueva de Callao continúan, por lo que esperamos la recuperación de nuevos restos nos ayuden a despejar las dudas que esta nueva especie plantea.
Siete dientes maxilares (designados desde CCH6-a a CCH6-e, CCH8 y CCH9), dos falanges de la mano (CCH2 y CCH5), dos falanges del pie (CCH3 y CCH4) y un fémur fracturado (CCH7). ↩
Cinco de los dientes pertenecen a un mismo individuo (CCH6-a a CCH6-e). Por otro lado, la presencia de dos terceros molares superiores derechos (M3; CCH6-a y CCH9) confirman otro; mientras que el fragmento del fémur que pertenece a un juvenil (CCH7) representa al tercero. ↩