Última actualizacón: 6 julio 2018 a las 08:52

En esta nueva sección pretendo destacar los avances científicos que se han producido en la semana que termina, con enlaces directos a las noticias más relevantes e incluyendo los artículos originales para que el lector pueda acudir directamente a la fuente para tener una información más completa.
ECOLOGÍA
Esta semana hemos sabido que, según las últimas investigaciones, el Mar Mediterráneo ha perdido en el último medio siglo casi el 40 por ciento de la Posidonia oceánica, un ecosistema milenario dominante en este mar. Esta disminución tiene efectos en la cantidad de dióxido de carbono que este ecosistema captura que se estima entre el 62% y el 87 % del que secuestraba antes de 1960. Como era de esperar, el 67 % del declive se debe a impactos causados por el ser humano.
“La Posidonia oceanica, angiosperma marina endémica del Mediterráneo, forma el ecosistema costero dominante en este mar y proporciona importantes servicios: las praderas son sumideros de carbono, estabilizan el sedimento, evitan la erosión costera, e incrementan la biodiversidad y recursos vivos”, señala la investigadora del CSIC Núria Marbà.
La investigadora señala que este trabajo “ha evaluado el estado (estable, en expansión o en regresión) generalizado de las praderas de Posidonia oceanica en el Mediterráneo desde que existen medidas (1842) de extensión, cobertura y densidad de biomasa. Hemos obtenido datos de 519 praderas, el 97 % de ellas en la costa europea del Mediterráneo occidental”.
• Noticia Tendencias21
• Artículo: Mediterranean seagrass (posidonia oceanica) loss between 1842 and 2009
EVOLUCIÓN HUMANA
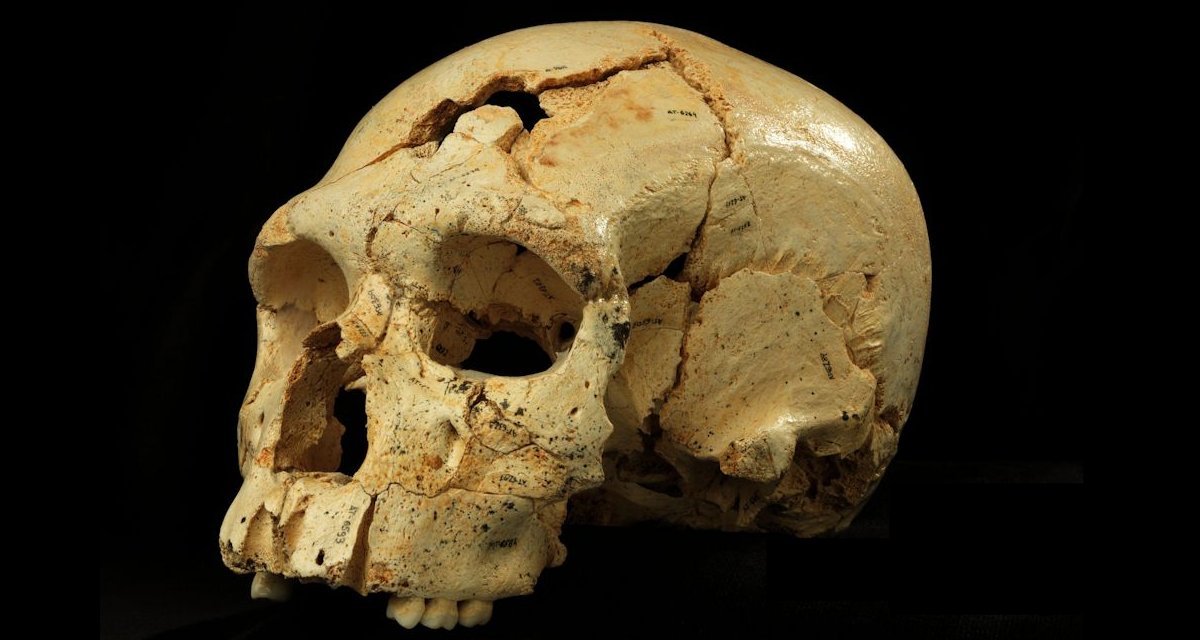
Diecisiete cráneos de homínidos, siete de ellos nuevos para la ciencia, acaban de aclarar cómo y cuándo comenzaron a surgir los rasgos característicos del rostro de los neandertales, esos primos de los humanos modernos que se extinguieron hace unos 30.000 años. Los restos se han hallado en la Sima de los Huesos de Atapuerca (Burgos), el mayor yacimiento de fósiles de homínidos de todo el mundo. Además muestra que los homínidos de la Sima de los Huesos, cuyos fósiles han sido datados de nuevo en unos 430.000 años, no pertenecen a la especie que se creía (Homo heidelbergensis) ni tampoco a la neandertal. Son una especie de punto medio entre ambos, aún sin nombre y que supone un nuevo revolcón para las teorías vigentes sobre evolución humana.
Un nuevo estudio asegura que los homínidos de la Sima de los Huesos han sido erróneamente identificados. Se trataría de los “primos abuelos” de los neandertales y ya tenían esas cejas prominentes y caras triangulares que caracterizaron a la especie
El estudio muestra que los rasgos neandertales comenzaron a aparecer progresivamente y que los primeros cambios se dieron en la cara y las mandíbulas. Solo hace falta palparse la cara para reconocer algunos de ellos, señala Martínez. Los neandertales tenían el arco de las cejas muy pronunciado y este no se hundía al pasar sobre la nariz, como sí nos pasa a los Homo sapiens. También, en nuestra especie, los pómulos se proyectan hacia afuera. Los de los neandertales estaban inclinados hacia la nariz, lo que les hacía tener una cara más triangular. Además su mandíbula tenía un encaje diferente con el cráneo.
El estudio afirma que los neandertales no aparecieron por un proceso lineal y con un solo grupo, sino en una serie de cambios que se produjeron en diferentes lugares y grupos, formando una especie de mosaico por toda Europa.
El equipo de Atapuerca aduce que en 1993, con muchos menos fósiles, atribuyeron a los homínidos de la Sima a heidelbergensis por falta de más datos. El Homo heidelbergensis es una especie descrita en 1907 en base a una mandíbula hallada en Alemania, con rasgos más primitivos que los de los homínidos de la Sima y que tiene unos 600.000 años. Con el paso de los años en Atapuerca se dieron cuenta de que los fósiles de la Sima no encajaban con los de heidelbergensis, pero ha tenido que pasar todo este tiempo hasta hacer una declaración “oficial y conjunta” con este estudio, reconoce Martinón-Torres.
Pero aún queda sitio para la polémica en el árbol genealógico de los humanos. Para Stringer, por ejemplo, heidelbergensis es nuestro ancestro, además de los neandertales. “Considero que heidelbergensis fue una especie con una gran expansión”, señala. “En África dio lugar a los Homo sapiens y en Europa y Asia dio lugar a los neandertales y sus parientes los denisovanos”, comenta.
En Atapuerca las cosas se ven diferentes. Dicen que fue el Homo antecessor, que vivió hace unos 1,3 millones de años en el mismo rincón de la sierra Burgalesa, el ancestro común entre sapiens y neandertales.
Y para el futuro próximo queda otra asignatura pendiente que puede levantar ampollas: ponerle nombre de especie, o no, a los homínidos de la Sima de los Huesos. “Si fueran fósiles de ratones o bisontes no habría duda de que esto es una nueva especie, pero al ser humanos hay mucha más atención y es necesario aportar muchos más datos”, concluye Martínez.
• Noticia Materia, El Mundo
• Artículo: Neandertal roots: Cranial and chronological evidence from Sima de los Huesos
MEDICINA
Investigadores de la Universidad de Cincinnati (UC), en Estados Unidos, han encontrado una terapia que invierte el comienzo de la diabetes tipo 1 en modelos de ratón y puede ayudar a avanzar en los esfuerzos para combatir la enfermedad entre los seres humanos.
En la diabetes tipo 1, el cuerpo no produce suficiente insulina, que es fundamental para el metabolismo de la glucosa, puesto que sin insulina, la glucosa en sangre se eleva, pero no existe una cura para esta patología, sólo se puede controlar con la terapia de insulina. Los síntomas de la enfermedad incluyen micción frecuente, sed excesiva y pérdida extraña de peso aunque se esté comiendo más. Los investigadores dicen que la incidencia de la diabetes tipo 1 y la autoinmunidad en general han aumentado rápidamente desde mediados del siglo XX, posiblemente como resultado de una menor estimulación del sistema inmunitario innato, lo que desencadena la autoinmunidad en niños y adultos jóvenes. En la diabetes tipo 1, la autoinmunidad hace que las células T del organismo ataquen a las células beta productoras de insulina. Anteriormente, se ha informado acerca de ratones diabéticos no obesos que tienen defectos en las células inmunes innatas y que TLR4, una proteína codificada por el gen TLR4, juega un papel protector en la prevención de la diabetes tipo 1.
Ridgway, profesor y director de la división de Inmunología, Alergia y Reumatología de la Universidad de California, en Estados Unidos, explica que su equipo de investigadores utilizó un anticuerpo monoclonal antagonista, UT18, para impulsar la actividad de TLR4 y revertir la diabetes de nueva aparición en un alto porcentaje de ratones no obesos diabéticos. «Hemos demostrado que mediante el uso de un anticuerpo para estimular una molécula específica en el sistema inmune innato podemos revertir, con una alta tasa de éxito, la diabetes de nuevo inicio en ratones que ya han comenzado a desarrollar síntomas de la enfermedad», subrayay Ridgway. «La causa de esta inversión es un mantenimiento de las células beta pancreáticas endocrinas que producen la insulina. Estas células se mantienen desde el ataque autoinmune, que es el sello distintivo de la diabetes tipo 1», añade.
La clave para revertir la diabetes tipo 1 en ratones, según Ridgway, es la captura de la enfermedad en su inicio, que se trata típicamente de una ventana de tiempo muy corto.
• Noticia La Razón
• Artículo: Reversal of New-Onset Diabetes in NOD Mice by an Agonist TLR-4/MD2 Monoclonal Antibody

Usando un tipo de célula madre humana, investigadores de la Universidad de Johns Hopkins (EEUU) han logrado crear en laboratorio una estructura tridimensional similar a la de la retina humana, que incluye células fotorreceptoras funcionales, capaces de responder a la luz. Los científicos consideran que este es un primer paso hacia la posibilidad de sustituir el tejido retiniano enfermo o muerto con material de laboratorio, desarrollado para restaurar la visión.
Al igual que muchos procesos del cuerpo, la visión depende de muchos tipos diferentes de células que trabajan concertadas, en este caso, para convertir la luz en algo que pueda ser reconocido por el cerebro como una imagen.
Canto Soler advierte que los fotorreceptores son sólo parte del proceso complejo de la visión que desarrollan los ojos y el cerebro, y que su laboratorio aún no ha recreado todas las funciones del ojo humano y su relación con la corteza visual cerebral.
«¿Es nuestra retina de laboratorio capaz de producir una señal visual que el cerebro pueda interpretar como una imagen? Probablemente no, pero este es un buen comienzo», señala.
El tejido de la retina es complejo, pues comprende siete tipos principales de células, entre ellos seis tipos de neuronas, que se organizan en capas de células específicas que absorben y procesan la luz, “ven”, y transmiten esas señales visuales al cerebro para su interpretación. Las retinas de laboratorio recrean esa arquitectura tridimensional de la retina humana.
«Sabíamos que una estructura celular en 3D era necesaria si queríamos reproducir las características funcionales de la retina», explica Canto Soler, «pero cuando comenzamos este trabajo, no pensamos que las células madre podrían acumularse una retina casi por su cuenta. En nuestro sistema, de alguna manera, las células supieron qué hacer».
Cuando el tejido de la retina de laboratorio se hallaba en una fase equivalente a las 28 semanas de desarrollo fetal en el útero, con los fotorreceptores ya bastante maduros, los investigadores probaron estas mini-retinas para ver si estos fotorreceptores podían de hecho registrar y transformar la luz en señales visuales.
Lo hicieron mediante la colocación de un electrodo en una sola célula fotorreceptora y luego enviando un pulso de luz a esta célula, que hizo que esta reaccionara siguiendo un patrón bioquímico similar al del comportamiento de los fotorreceptores en personas expuestas a la luz.
En concreto, dice Canto Soler, los fotorreceptores cultivados en laboratorio respondieron a la luz del mismo modo que lo hacen los bastones de la retina. Las retinas humanas contienen dos tipos principales de células fotorreceptoras, llamadas bastones y conos. La inmensa mayoría de los fotorreceptores de los humanos son bastones, que permiten la visión en condiciones de poca luz. Las retinas cultivadas por el equipo de la Johns Hopkins también tenían sobre todo bastones.
• Noticia Tendencias21
• Artículo: Generation of three-dimensional retinal tissue with functional photoreceptors from human iPSCs
MICROBIOLOGÍA
Un grupo de investigadores de Canadá ha encontrado una bacteria resistente a nuestros más potentes antibióticos –lo que se conoce como “superbacterias”, o bacterias multirresistentes–, en unos calamares congelados de una tienda de alimentación china en Saskatoon, al sur del país, que según su propietario provenían de Corea del Sur.
Hasta ahora, tal como ha explicado a El Confidencial Álvaro Pascual Hernández, catedrático de microbiología de la Facultad de Medicina de la Universidad de Sevilla, no era extraño encontrar bacterias multirresistentes en la comida cruda, pero nunca se había encontrado una resistente a los Carbapenem, el tipo de antibióticos más potentes, que constituyen hasta el momento la última resistencia con la que contamos para combatir a las bacterias más peligrosas.
La mayoría de estas «superbacterias» habían aparecido hasta ahora en centros sanitarios, donde la presión de los medicamentos es muy grande, y habían sido trasmitidas a la población por pacientes de los centros. El último de estos brotes, en 2011, mató a siete personas en EEUU.
Pero el descubrimiento de la bacteria en una tienda de alimentación, tras un control rutinario, se ha hecho público en un informe del Centro para el Control y la Prevención de Enfermedades de Estados Unidos. Y ha encendido todas las alarmas.
Según ha explicado al Washington Post el autor del informe, Joseph Rubin, profesor de microbiología en la Universidad de Saskatchewa y miembro del equipo de investigadores que identificaron la bacteria, el descubrimiento de una “superbacteria” de este tipo en la comida implica que “el riesgo de exposición del público va más allá de los viajeros y la gente que ha sido hospitalizada. Significa que un segmento mucho más amplio de la población está en riesgo. Es algo que puedes traer a tu casa”.
La bacteria encontrada en los calamares es un organismo muy común, Pseudomonas fluorescens, presente en el suelo y el agua, y no es peligrosa. De hecho, al cocinar los calamares habría muerto. Pero el organismo podría haber infectado a cualquier persona gracias a la contaminación cruzada, muy habitual si no tomamos las debidas precauciones al cocinar y olvidamos lavarnos las manos o limpiar la tabla de cortar; o si se hubieran consumido los calamares crudos, algo que no es tan raro.
Tal como explica Pascual, el hallazgo no es una amenaza directa de salud, pero sí la constatación de que «tenemos un problema muy serio de resistencia a los antibióticos». El problema no es la bacteria en sí, que no es patógena, sino la enzima o gen que la hace resistente a nuestros más poderosos medicamentos, los Carbapenem, que constituyen la última línea de defensa contra los microbios.
• Noticia El Confidencial
• Artículo: Carbapenemase-producing Organism in Food, 2014
NEUROCIENCIA
Una nueva investigación, liderada por los investigadores de la Universidad de Southampton, ha descubierto la neurogénesis, es decir, el mecanismo de reparación automática del cerebro adulto que puede ayudar a preservar la función del cerebro en las enfermedades neurodegenerativas como el Alzheimer, priones (trastorno neurodegenerativo) o el Parkinson.
La degeneración progresiva y la muerte del cerebro, que ocurre en muchas enfermedades neurodegenerativas, muchas veces visto como un proceso imparable e irrevocable. Sin embargo, el cerebro tiene un potencial autorreparador que es fundamental para la renovación de ciertas poblaciones de neuronas en el cerebro.
En el pasado la neurogénesis se había visto en las enfermedades neurodegenerativas. Sin embargo ahora un equipo de investigación conducido por el Dr. Diego Goméz-Nicola del centro de ciencias biológicas de la Universidad de Southampton, ha detectado un aumento de la neurogénesis en el giro dentado que contrarresta parcialmente la pérdida neuronal (foto 1) que se produce en enfermedades como el Alzheimer.
Se ha utilizado un modelo de enfermedad priónica (proteína que produce enfermedades neuronales degenerativas como podría ser el Alzheimer) en ratones y la investigación identificó el tiempo en que se producía la generación de estas neuronas recién nacidas y lo integró en los circuitos del cerebro. Se observó que mientras este mecanismo de reparación automática es eficaz para mantener algunas funciones neuronales al principio y a mitad de esta etapa, falla en las etapas más avanzadas de la enfermedad. Este avance supone una ventana temporal para la intervención terapéutica potencial, con el fin de preservar los efectos beneficiosos de la neurogénesis mejorada.
El Dr. Goméz-Nicola ha dicho que “este estudio pone en evidencia que el cerebro tiene la capacidad de poder orquestar una respuesta autorreparadora. La continuación de esta línea de investigación está abriendo nuevos caminos para identificar qué señales específicas se utilizan para promover esta mayor respuesta neurogénica, y centrarse más en la neurogénesis como enfoque terapéutico para promover la regeneración de las neuronas perdidas”.
• Noticia Ciencia y biología
• Artículo: Temporal dynamics of hippocampal neurogenesis in chronic neurodegeneration (Acceso directo en PDF)
FÍSICA
Al contrario de lo que ocurre con otras partículas elementales, los quarks no se observan aislados en la naturaleza. Aparecen siempre agrupados en forma de hadrones; es decir, en partículas como el protón, el neutrón o el pion. Hasta hace poco solo se conocían dos grandes clases de hadrones: bariones, o partículas formadas por tres quarks (como el protón), y mesones, compuestos por dos.
Ahora, en un artículo publicado hace unos días en Physical Review Letters, los investigadores del experimento LHCb del CERN han referido la existencia de una nueva partícula compuesta por cuatro quarks. Bautizada como Z(4430)–, el nuevo objeto tiene una masa de unos 4475 megaelectronvoltios (unas cuatro veces y media más masivo que el protón), posee espín 1 y su estructura puede describirse en términos de un estado ligado de los quarks c, anti-c, d y anti-u.
En 2008 el experimento japonés Belle obtuvo los primeros indicios sobre la existencia de esta partícula. Sin embargo, sus propiedades y estructura estaban aún pendientes de confirmarse. Ahora, gracias al elevado número de eventos analizados por la colaboración LHCb, los investigadores afirman haber eliminado otras interpretaciones más prosaicas del nuevo estado, como que se tratase de una «molécula» de mesones D. Según señalan en su artículo, el nuevo análisis deja el estado ligado de cuatro quarks «como única explicación plausible» de los datos.
Hace un año, los experimentos BESIII, en China, y Belle hallaron indicios de la existencia de otro tetraquark, al que apodaron Zc(3900). La confirmación empírica de esta nueva clase de estados hadrónicos tal vez permita a los investigadores profundizar en las propiedades matemáticas de la cromodinámica cuántica, la teoría que describe las interacciones entre quarks. Aunque sus ecuaciones básicas se conocen desde hace cuarenta años, sus predicciones a bajas energías no se dejan describir mediante las técnicas de cálculo habituales, basadas en diagramas de Feynman.
• Noticia IyC
• Artículo: Observation of the Resonant Character of the Z(4430)− State
GEOLOGÍA

Dos estudios diferentes han demostrado que el movimiento de las placas tectónicas (y por lo tanto de la corteza terrestre) no es constante, sino que varía con el transcurso del tiempo. Lo cual puede llevar a explicar los patrones de movimiento continental observados por los científicos en todo el mundo y, por lo tanto, también sus implicaciones a la hora de elaborar modelos climáticos.
Ambos trabajos han sido presentados hace unos días en el transcurso de la Conferencia Goldschmidt 2014, la más importante del mundo en el campo de la geoquímica, celebrada en la localidad californiana de Sacramento.
Un grupo de investigadores dirigido por Peter Cawood, de la universidad británica de St Andrews, ha analizado con detalle numerosas medidas del movimiento de los continentes y los procesos geológicos descritos en estudios previos. Los resultados indican que durante el periodo que va entre hace 1.700 millones de años y 750 millones de años, la Tierra pareció atravesar una larga era de estabilidad, en términos medioambientales, con muy poca construcción de nueva corteza y escasas fluctuaciones en la composición de la atmósfera, lo cual también contribuyó a la estabilidad de los seres vivos de aquella época.
Pero este periodo de mil millones de años de tranquilidad se contradice de forma muy marcada con lo sucedido en los periodos inmediatamente anterior y posterior a ese lapso de tiempo. De hecho, tanto antes como después se produjeron grandes edades de hielo y cambios bruscos en los niveles de oxígeno atmosférico. Los mil millones de años citados coinciden también con la formación del supercontinente Rodinia, que parece haber permanecido estable durante todo ese tiempo.
Peter Cawood sugiere que dicha estabilidad podría deberse al enfriamiento gradual de la corteza terrestre a lo largo del tiempo. «Antes de hace 1.700 millones de años -explica- la corteza terrestre podría haber estado sustancialmente más caliente, lo cual implica que el movimiento de los continentes estuvo entonces gobernado por reglas muy distintas a las que rigen en la actualidad.
Al final del periodo, hace 750 millones de años, la corteza se había ya enfriado lo suficiente como para permitir que la moderna tectónica de placas empezara a funcionar, y pusiera en marcha, como hace en la actualidad, el proceso de formación de zonas de subducción (en las que una placa se desliza debajo de otra). Este aumento de actividad podría haber desencadenado, a su vez, una miriada de cambios, incluida la rotura de Rodinia y la alteración de los niveles de elementos clave en la atmósfera y en los mares, lo que hizo posible que se produjeran los cambios evolutivos que permitieron el desarrollo de las formas de vida que vemos en el presente».
• Noticia ABC
• Artículo: Earth’s middle age