
Última actualizacón: 24 septiembre 2017 a las 13:05
Creo que todos conocemos la historia que voy a resumir: dos horas después de la medianoche del 12 de octubre de 1492, Rodrigo de Triana gritó desde La Pinta «¡Tierra a la vista!», poniendo fin de esta manera a una larga travesía por el Océano Atlántico. La expedición española, capitaneada por Cristóbal Colón, había llegado a la isla de Guanahaní, perteneciente al archipiélago de Las Bahamas. A la mañana siguiente Colón desembarcaba acompañado de Martín Alonso y Vicente Yáñez Pinzón, capitanes de La Pinta y La Niña respectivamente, y tomaba posesión de la tierra descubierta en nombre de la Corona de Castilla. Colón, desde ese momento, pasaría a la historia como el descubridor de América.

Colón desembarca en la isla de Guanahaní
La trascendencia de ese momento llega hasta nuestros días ya que el 12 de octubre se ha escogido como efeméride para la celebración de la Fiesta Nacional de España (Día de la Hispanidad, de Cristóbal Colón y otros tantos apelativos en diferentes países hispanohablantes), como símbolo del momento en que se inició el contacto entre Europa y América y que culminó con el llamado «encuentro de dos mundos». Sin embargo, hay quienes no están de acuerdo con esta versión de los hechos.
Más de cuatro siglos después de ese desembarco histórico, concretamente el 11 de octubre de 1965, se organizó una fiesta en la Universidad de Yale, en New Haven, Connecticut. Asistieron como invitados especiales un numeroso grupo de diplomáticos, universitarios y periodistas escandinavos a quienes se hizo partícipes en primicia del que fue considerado el descubrimiento cartográfico más fascinante del siglo: un mapa sobre pergamino de color beige de 27,8 centímetros de alto por 40 centímetros de largo —bautizado como el mapa de Vinlandia— que se afirmó era el único ejemplo de cartografía escandinava medieval que existía 1. Según los expertos que analizaron el documento, el mapa era auténtico y constataba que los vikingos —encabezados por Leif Eriksson— habían descubierto, explorado y colonizado una tierra fértil al oeste de Groenlandia en el siglo XI. Y además habían dejado constancia escrita de su hazaña. Por tanto, sostenían que ese mapa era la prueba palpable de que éstos se habían adelantado a Cristóbal Colón en más de tres siglos 2.
El mapa de Vinlandia se dio a conocer al público en general cuando la editorial universitaria de Yale lazó la obra The Vinland map and the Tartar relation 3 de forma simultánea a ambos lados del Atlántico (en nueva York y Londres); una monografía que pronto se convirtió en un best seller internacional.
Los periódicos de medio mundo se hicieron eco de la noticia y las reacciones ante este hallazgo no se hicieron esperar. Aunque de interés intrínseco para comprender mejor la historia de los tártaros (por el texto que lo acompañaba), el mapa no hubiera causado tanta sensación si no fuera por la descripción de una tierra situada en el noroeste del Océano Atlántico y al oeste de Groenlandia llamada Vinlandia. Igual de llamativo era que Groenlandia apareciera como una isla, cuando este hecho no se verificó hasta 1912 cuando se circunnavegó por primera vez.

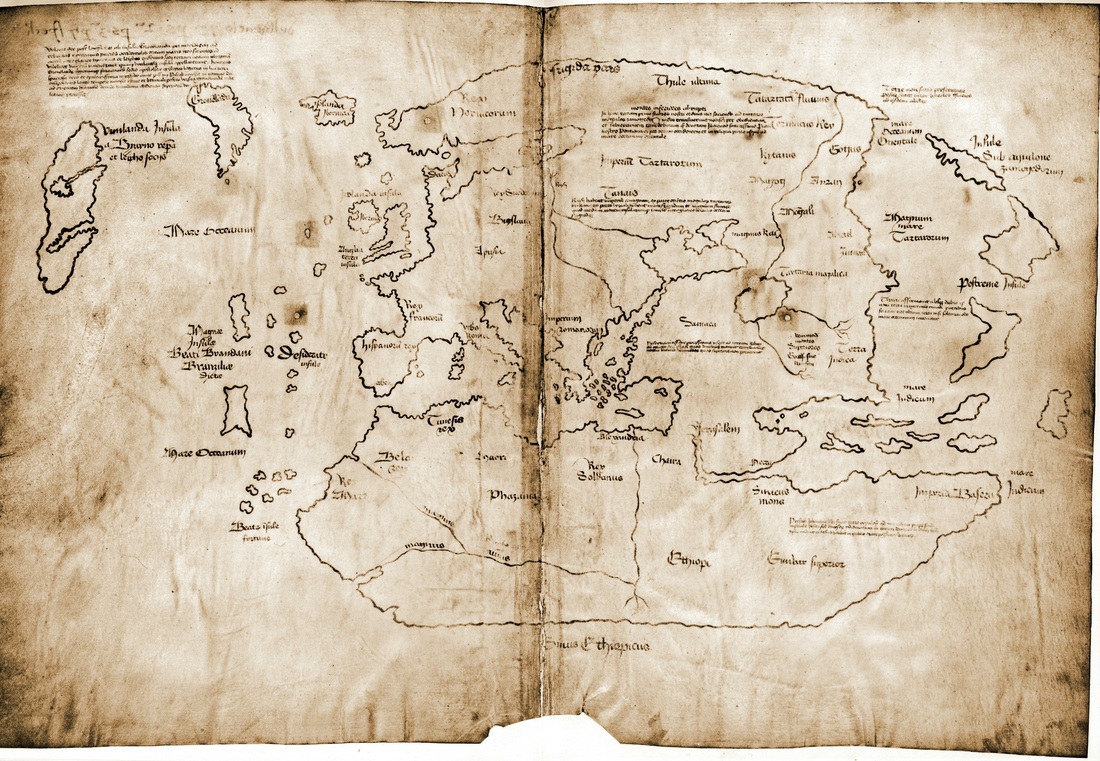
Mapa de Vinlandia (falso color)
En cualquier caso, para quienes recibieron la noticia no pasó desapercibida la fecha escogida para dar publicidad al hallazgo: la víspera de la conmemoración del descubrimiento “oficial” de América por Cristóbal Colón. En este sentido, y con un tono que desborda ironía, Torcuato Luca de Tena analizó lo sucedido en un artículo publicado en ABC dos días después 4. Además de poner en duda la autenticidad del mapa, criticó duramente que por parte de las autoridades académicas de la universidad americana se hubiera escogido precisamente ese día, considerando que se trató de una cuestión política más que científica:
La gesta escandinava producirá una huella interesante en el campo de las artes. Va a ser preciso arrinconar los cuadros de las carabelas colombinas de Rodrigo de Triana, empinado sobre la cofa, y de Colón, con el mandoble desenvainado, mientras se remoja los pies al desembarcar en el hemisferio de Ericsson. Las puertas de bronce del Capitolio de Washington recogen la escena de Colón cuando estaba en el lecho de muerte. Habrá que enterrar pronto a este enfermo, para poner a Ericsson en la misma cama. […] Gracias a ese reajuste muy pronto se podrá hablar de las grandes universidades de Vinlandia, de las famosas aguas de Vinlandia a base de colas, y del Presidente Johnson de los Estados Unidos de Vinlandia.
Cuando supe que Scientia, anfitrión de la décimo primera edición del carnaval de humanidades, había elegido como tema central el arte y la química, me vino a la mente de inmediato la historia de este mapa, que quizás muchos ya conozcan, pero que aún guarda interesantes incógnitas que vamos a analizar en esta serie de anotaciones. Para mí resulta un tema apasionante porque combina algunas de las cuestiones que más me atraen: historia, navegación, arqueología, manuscritos antiguos y un enfrentamiento científico que ha durado varias décadas (y que aún sigue vivo como vamos a comprobar) a cuenta de los numerosos análisis químicos realizados sobre el mapa.
El mapa de Vinlandia
El primer trabajo de estudio y análisis de los documentos (mapa de Vinlandia y Relación tártara) fue llevado a cabo como hemos apuntado por el Dr. R. A. Skelton, superintendente de la sala de mapas del Museo Británico; el Dr. Thomas E. Marston, conservador de literatura medieval y renacentista de la biblioteca de la Universidad de Yale; y George D. Painter, ayudante de conservador a cargo de los incunables del Museo Británico.
Según refieren estos investigadores en la monografía publicada en 1965, el mapa fue elaborado alrededor del año 1440 por un escriba desconocido, empleando como modelo un original de fecha anterior del que no se tiene constancia. Junto al mapa apareció una versión manuscrita —también desconocida hasta entonces— 5 del informe del viaje que el misionero franciscano de origen italiano Giovanni da Pian del Carpine hizo a la corte mongola en 1245 (texto que por ese motivo recibe el nombre de Relación tártara) escrita por un fraile llamado C. de Bridia, y cuyo nombre tampoco aparece en ningún registro histórico. La Relación tártara es un texto breve que ofrece una versión algo diferente de la bien conocida Ystoria Mongalorum escrita por Carpine, jefe de la misión enviada por el Papa Inocencio IV al rey de los tártaros. La expedición dejó Lion en abril de 1245 y estuvo fuera durante dos años y medio. Se trata de uno de los varios informes que describen los viajes de los misioneros durante el siglo XIII a Asia Central así como la historia y naturaleza de los mongoles.
Un año después, la misma universidad compró otro manuscrito, el Speculum historiale de Vincent de Beauvais, la tercera parte de una obra mayor que pretendía ser un relato histórico del devenir del mundo. La importancia fundamental de esta última adquisición proviene del hecho de que, según sostienen los investigadores, las tres piezas formaban parte de un mismo libro y fueron redactadas por una misma mano (más adelante ahondaremos en esta cuestión). Por lo tanto, afirman que el Speculum fue separado físicamente de un volumen donde el mapa se situaba al principio y la Relación tártara al final.
Describamos el mapa. Está dibujado en una hoja de pergamino doblada por la mitad y contiene, en líneas generales, una descripción de las tres partes conocidas en el mundo medieval —Europa, África y Asia— rodeadas por el océano, con islas y grupos de islas tanto en el este como en el oeste. El mapa se ha dibujado con orientación norte, es decir, con los nombres y referencias escritos de forma horizontal en una alineación este-oeste del territorio (una cuestión nada baladí porque no era lo habitual en la época. De hecho se desconoce si esta disposición fue deliberada o suponía el método más conveniente para ordenar los elementos del mapa dentro del espacio rectangular del que disponía el escriba).
En el contorno de las costas se representan las características hidrológicas (ríos y mares interiores) aunque los detalles orográficos no aparecen reflejados. El dibujo se ha hecho con una línea de tinta gruesa, y muestra una generalización evidente en algunas partes y una considerable elaboración más detallada en otras.
Lo más relevante a efectos de esta anotación es que en el extremo noroeste del mapa aparecen tres grandes islas, llamadas respectivamente isolanda Ibernica, Gronelada, y Vinlanda Insula a Byarno repa et leipho socijs —es decir, Islandia, Groenlandia y Vinlandia—. El hecho de que estas islas aparezcan fuera del marco oval del resto del mapa sugiere que no estaban en el modelo original que el cartógrafo siguió al copiar el resto. En la esquina superior izquierda se incluye una larga referencia al viaje del obispo Erik Gnupsson a Vinlandia:
Por la voluntad de Dios, después de un largo viaje desde la isla de Groenlandia hacia las partes más distantes del mar océano occidental, navegando hacia el sur, en medio del hielo, los compañeros Bjarni [Herjolfsson] y Leif Eriksson descubrieron una nueva tierra, extremadamente fértil, que incluso tenía viñas, a la que llamaron Vinlandia. Erik, legado de la Sede Apostólica y obispo de Groenlandia y las regiones vecinas, llegó a esta tierra verdaderamente inmensa y muy rica, en el nombre de Dios Todopoderoso, en el último año de nuestro bienaventurado padre Pascal, permaneció mucho tiempo, tanto en verano e invierno, y más tarde regresó hacia el noreste hacia Groenlandia y luego se dirigió en la mayor humilde obediencia a la voluntad de sus superiores.

Detalle del mapa de Vinlandia (esquina superior izquierda)
Para los investigadores, la asociación física del mapa con el resto de manuscritos quedaba demostrada más allá de toda duda por tres pares de agujeros causados por gusanos y que atraviesan sus dos hojas. Éstos coinciden perfectamente con los que se encuentran en las hojas de comienzo del Speculum, coincidiendo a su vez los de éste con los que presenta la Relación tártara. De ahí que el hallazgo de la obra de Beauvais fuera tan importante: era la pieza que otorgaba autenticidad a todo el conjunto.
¿De dónde ha salido el mapa?
Desde el mismo momento en que se hizo público el hallazgo de este singular documento cartográfico, tanto periodistas como estudiosos de diferentes nacionalidades pusieron en tela de juicio su autenticidad.
Uno de los principales problemas a los que tuvo que enfrentarse la Universidad de Yale tras su adquisición fue la falta de un registro ininterrumpido de su posesión, es decir, no había un conocimiento detallado de sus anteriores propietarios y las incidencias por las que habían atravesado los textos. Skelton, Marston y Painter (que, recordemos, llevaron el peso de la investigación incial) reconocieron desde el principio que sin ese registro no podía haber una absoluta e irrefutable demostración de que el mapa no era una falsificación 6. Conviene por tanto conocer cómo apareció el mapa.
La historia del volumen se conoce desde el verano de 1957 cuando Enzo Ferrajoli de Ry, un librero italiano afincado en Barcelona, lo dio a conocer en el reducido mundillo del mercado de libros raros. En Londres, acompañado por el librero Joseph Irving Davis, de Davis y Orioli Ltd., Ferrajoli se lo ofreció al Museo Británico, donde George Painter, el Dr. Skelton y el Dr. Schofield pudieron echarle un vistazo. Éstos apreciaron de inmediato las características novedosas del mapa aunque, en ese momento, expresaron serias dudas acerca de algunas inscripciones latinas que contenían aparentemente algunos “errores monumentales”. La venta no se materalizó.
Continuando con su periplo, en septiembre de ese mismo año tenemos a Ferrajoli en la oficina que Nicholas Rauch tenía en Ginebra, donde el anticuario de New Haven Laurence Claiborne Witten II vio por primera vez tanto el mapa como de la Relación tártara. Esta vez nuestro barcelonés de acogida sí tuvo éxito. Más tarde, en la Conferencia sobre el mapa de Vinlandia celebrada en noviembre de 1966 en la Smithsonian Institution (de la que hablaremos luego), el propio Witten rehusó revelar la procedencia de los manuscritos más allá de indicar que eran propiedad de una familia en cuya biblioteca habían permanecido durante un par de generaciones y que él visitó personalmente. Reconoció haber pagado 3.500 dólares al propietario por el volumen que contenía el mapa de Vinlandia y la Relación tártara, abonando a Ferrajoli una comisión por su participación. Esto es lo que mantuvo Witten cuando, de forma insistente, se le pidió que aclarase cómo supo de la existencia del mapa aunque, como veremos al final, hay otra versión más plausible. Baste que sepamos esto por ahora.
Finalmente, con Witten ya de vuelta en Estados Unidos en octubre de 1957, entran en escena Thomas Marston y Alexander Vietor, conservador de literatura medieval y renacentista, y conservador de mapas respectivamente, trabajando ambos en la biblioteca de la Universidad de Yale. Witten les llevó los documentos con la intención de que la biblioteca los adquiriese aunque, cuando Marston y Vietor vieron el mapa, no pudieron verificar su autenticidad: se dieron cuenta de que la encuadernación era moderna (del siglo XIX) mientras que el texto de la Relación tártara era del siglo XV. Además, se suponía que el mapa era una ilustración de la Relación pero los agujeros producidos por los gusanos no coincidían. Por último, había una inscripción extraña en el reverso del mapa: “Descripción de la primera parte, segunda parte (y) tercera parte del Speculum”. Esta anotación se refería por tanto a un libro diferente a la Relación tártara. En cualquier caso, la Universidad de Yale no disponía de medios económicos para su compra así que Witten regaló a su esposa el manuscrito.
Éste estaba aún investigando la autenticidad del documento cuando Marston le encargó a principios de 1958 que adquiriera del catálogo de Davis y Orioli (recordemos, la empresa de libreros británica que intervino en el intento de venta de los textos al Museo Británico) un manuscrito con parte del trabajo de un monje dominico del siglo XIII llamado Vincent de Beauvais: se trataba del Speculum historiale, por el que pedían 75 libras. Cuando Witten sostuvo en sus manos el libro tuvo una intuición que más tarde se confirmó, era la pieza que faltaba en la composición original: el mapa de Vinlandia al principio, el Speculum en medio, y la Relación tártara al final. Las marcas de agua en el papel de los dos libros eran las mismas 7 y permitieron situar su fecha de origen en 1440, probablemente en la ciudad suiza de Basilea. Tanto el Speculum como la Relación tártara parecían el trabajo del mismo escriba, quien empleaba en ambos la misma caligrafía conocida como “Upper Rhineland bastard» (o cursiva del Alto Rin). Pero lo que entusiasmó a Marston y Witten fue que la letra parecía la misma también en el mapa.
El Dr. Marston, después de mucho pensarlo, regaló el Speculum a la Sra. Witten aunque ahora los tres pergaminos se habían vuelto enormemente valiosos, económica e históricamente. Marston esperaba que ese gesto de generosidad le daría a la biblioteca de Yale alguna posibilidad de control sobre el mapa en el caso de que la Sra. Witten decidiera venderlo, como finalmente sucedió.
De esta forma, el mapa fue adquirido por Paul Mellon, antiguo alumno de la universidad, quien accedió a pagar lo que pedía Witten para donarlo a la biblioteca de Yale si podía ser autentificado. Reconociendo su importancia potencial como el mapa más antiguo en mostrar América, Mellon insistió en que su existencia se mantuviera en secreto hasta la publicación de un libro donde se estudiase y analizase en profundidad. Incluso los tres autores que debían escribir la obra fueron escogidos entre el pequeño número de personas que habían visto el mapa antes que él. Una vez publicado en 1965, cumpliendo su compromiso, lo donó a la Biblioteca Beinecke de Libros Raros y Manuscritos de la Universidad de Yale, donde se custodia en la actualidad.

Primer folio de la Relación tartara
Primeros análisis
Debemos tener en cuenta que es difícil que una investigación científica aporte una prueba definitiva sobre la autenticidad de un objeto artístico antiguo, salvo que sea posible demostrar de forma directa el momento de su fabricación. Así sucede, por ejemplo, cuando se emplea la técnica de datación por termoluminiscencia a la cerámica recuperada en yacimientos arqueológicos. Por tanto, la prueba casi perfecta de la autenticidad de un objeto es disponer de un registro de custodia ininterrumpida desde una conocida y probada fecha de origen (algo casi imposible en la mayoría de los casos).
Por suerte, cualquier documento, manuscrito o texto impreso, presenta una serie de componentes que sí pueden ser objeto de un detallado análisis: el papel o pergamino; la tinta; la encuadernación (en el caso de un libro) y otros aspectos accidentales como pueden ser la actividad de determinados insectos, las manchas etc. Cada uno de estos componentes puede someterse en mayor o menor medida a un examen científico con el fin de determinar la identidad o la forma de preparación de los mismos y cuyos resultados pueden ser comparados con muestras conocidas para proceder a su autentificación.
En la monografía publicada en 1965, los autores expusieron que se habían realizado todas las pruebas que no implicaban causar daños o destruir el pergamino (lo que en términos prácticos, y teniendo en cuenta la tecnología disponible en ese momento, no era demasiado). De hecho, reconocían sentirse frustrados porque aún no se hubiera perfeccionado algún tipo de análisis químico que sirviera para determinar tanto la antigüedad como el punto de origen aproximado del pergamino, ya que las que existían en ese momento no eran lo suficientemente precisas. Tampoco se hicieron análisis químicos de la tinta ya que era necesario raspar una gran cantidad de escritura para producir suficiente material con el que trabajar 8.
Sin embargo, pasaron por alto —o se negaron a hacer— unas pruebas bastante sencillas y no invasivas sobre la tinta utilizada en los textos que hubieran arrojado luz acerca de la autenticidad del mapa de Vinlandia.
Las tintas más utilizadas en la Edad Media para escribir sobre pergamino eran las metaloácidas o ferrogálicas dado su poder de fijación sobre ese soporte 9. La tinta ferrogálica está compuesta por cuatro ingredientes básicos: extractos de ácidos gálico y tánico (obtenido de las nueces de agalla del roble), vitriolo (minerales como sulfato ferroso o sal de hierro), y como aglutinantes goma arábiga y agua. La tinta fabricada según esta “receta” era transparente, lo que obligaba a añadir una pequeña cantidad de carbón en polvo (hollín) para guiarse durante la escritura. De esta forma, a medida que se iba secando el agua, los metales reaccionaban con los ácidos, provocando la oxidación y el oscurecimiento del compuesto que formaba un tanino férrico. El color original de estas tintas era azul intenso, casi negro, pero dado que la oxidación que sufren los metales es progresiva, el paso del tiempo produce alteraciones químicas que generan distintas variaciones cromáticas que van desde el pardo oscuro hasta el anaranjado.
Cuando el estado de corrosión es evidente, las características más significativas de la tinta ferrogálica son su difusión en el soporte, pérdida de nitidez y una apariencia quemada. Los ácidos presentes en estas tintas causan la “migración” de la escritura de una página a otra, o que se transfiera hasta aparecer visiblemente en el reverso del documento. Por lo tanto, el mero paso del tiempo (proceso que puede acelerarse o retardarse en función del cuidado que se dispense en la conservación) permite que la corrosión sea identificada a simple vista, ya que produce una apariencia de manchas borrosas y sombras.
Así, tras un examen no invasivo, los investigadores concluyeron que la caligrafía y la naturaleza física de los materiales empleados (tanto el pergamino, como el papel y la tinta) eran compatibles con la fecha que se había fijado para la fabricación del soporte físico: el año 1440, décadas antes de que Colón emprendiese su viaje de descubrimiento.

¿Los vikingos descubrieron América?
Tras el revuelo causado por la publicación del mapa, el 24 de febrero de 1966 se celebró una reunión privada de la Sociedad de Anticuarios británica donde Skelton leyó una nota en la que, tras escuchar las críticas que habían planteado algunos especialistas, reconoció que era necesario llevar a cabo un minucioso análisis químico de la tinta antes de poder aceptar la premisa de que tanto el manuscrito como el mapa habían sido confeccionados por la misma persona. En cambio, Marston defendió que los análisis químicos no aportarían nada nuevo, insistiendo en que no merecía la pena realizarlos dado el posible daño que se causaría al manuscrito.
En cualquier caso, las críticas no se ciñeron únicamente al poco peso de las pruebas aportadas que pretendían determinar la antigüedad de los materiales empleados. G. R. Crone, bibliotecario y conservador de mapas de la Real Sociedad Geográfica británica, planteó cuestiones estrictamente cartográficas 10, al igual que la doctora Eva Taylor, quizás la mayor autoridad mundial en mapas medievales de la época. Taylor pudo ver una reproducción del mapa en 1962 mientras Skelton trabajaba en él, y tras un examen largo y minucioso, elaboró una lista de objeciones de las que le hizo partícipe aunque finalmente ninguna fue tomada en consideración por el equipo de Yale. Por ese motivo preparó un artículo para ser publicado tras la aparición del libro donde ofrecería su versión del mapa 11. La Dra. Taylor mantenía que el mapa no podía ser un producto cartográfico del siglo XV porque dibujaba el contorno de Groenlandia como una isla y con gran detalle, algo que no se supo hasta siglos después (sus palabras textuales fueron que «colocado [el mapa de Vinlandia] junto a un mapa del siglo XX de una escala aproximadamente igual, ambos resultan casi indistinguibles a primera vista»). Otros puntos conflictivos eran la situación equivocada de Creta, el dibujo incorrecto del mar Egeo y la ausencia del mar de Mármara, todo en una época en la que ya existían mapas detallados del Mediterráneo que sí recogían todas estas características.
A pesar de todo, el mapa seguía levantando una importante expectación. A comienzos de 1967 los manuscritos comenzaron una gira europea para ser exhibidos en Noruega, Holanda y Gran Bretaña. Tanto el mapa como la Relación tártara permanecieron un par de días en el laboratorio de investigación del Museo Británico para someterlo a un estudio comparativo de la escritura mediante el empleo de métodos ópticos sencillos: iluminación ultravioleta y microscopía de baja potencia. Los resultados constataron que no había hierro en la tinta del mapa de Vinlandia, un hecho muy extraño para la época en que supuestamente fue dibujado. Por ese motivo se llevó a cabo una búsqueda sistemática de manuscritos del siglo XV que no emplearan hierro entre los componentes de sus tintas. No hubo éxito en dicha búsqueda.
Este fue uno de los factores que llevó a la Universidad de Yale —presionada por el siempre escéptico Departamento de Historia de la institución— a poner los manuscritos y el mapa en manos del químico norteamericano Walter McCrone. Su laboratorio debía realizar un análisis de las tintas utilizadas en el mapa, en el Speculum historiale y la Relación tártara. Para ello se tomaron distintas muestras, aunque no fue hasta 1972 cuando se contó con las herramientas y técnicas de microanálisis adecuadas para el trabajo. Los resultados se hicieron públicos el 26 de enero de 1974 12 y cayeron como un jarro de agua fría: el mapa parecía ser una falsificación realizada en algún momento posterior a 1920, mientras que el Speculum y la Relación tártara parecían ser genuinamente antiguos.
Continúa… con la segunda parte.
![]()
Esta entrada participa en la XI Edición del Carnaval de Humanidades, cuyo blog anfitrión es SCIENTIA.
Esta entrada participa en el XXXVIII Carnaval de la Química cuyo blog anfitrión es Pero esa es otra historia…
![]()
Notas
- Los vikingos eran tan buenos navegantes que no necesitaban cartas, se bastaban con seguir las estrellas. ↩
- Tres años después de que el mapa de Vinlandia viera la luz, el descubrimiento llevado a cabo por un equipo de arqueólogos noruegos de unas ruinas escandinavas del siglo XI en Terranova reforzó esta teoría. Para algunos, las ruinas representan la prueba física de que navegantes vikingos habían surcado el Atlántico Norte desde Islandia y Groenlandia hacia el oeste. Volveremos sobre este tema más adelante. ↩
- Skelton, R. A.; Marston, T. E. y Painter, G. D. (1965), The Vinland map and the Tartar relation. New Haven: Yale University Press, xii, 291 p. ↩
- En la última parte de esta serie he incluido algunos recortes de periódicos de la época que ofrecen una visión fresca del revuelo mediático que desde entonces ha tenido todo lo relacionado con el mapa. ↩
- Todo en una encuadernación del siglo XIX. ↩
- En cualquier caso, añadieron que el análisis del contenido y la forma en el contexto histórico puede crear —y, en este caso, estaban convencidos de que había creado— una presunción de autenticidad que era muy difícil, si no imposible, discutir. ↩
- Aunque la historiadora Kirsten Seaver ha señalado que los sellos presentes en páginas aleatorias del libro indican una propiedad institucional, no privada, algo sobre lo que volveremos luego. ↩
- Debido a la menor capacidad de absorción del pergamino sobre el papel, las tintas se depositan en la superficie sin llegar a penetrar entre sus fibras. ↩
- De hecho, es curiosa la etimología de la palabra tinta: deriva del latín encaustum o incaustum —quemado—, que son el origen del francés encre, del inglés ink, y del italiano inchiostro. El término podría provenir de la oxidación de los ácidos gálico y tánico (dos de los componentes de la tinta) que hace que se perfore o queme la superficie escrita. ↩
- Crone, G. R. (1966), «How authentic is the «Vinland Map»?». Encounter, núm. 26, p. 75-78. ↩
- Richey, M. W. (1966), «The Vinland map». The Journal of Navigation, vol. 19, núm. 01, p. 124-125. ↩
- McCrone, W. C. y McCrone, L. B. (1974), «The Vinland map ink». The Geographical Journal, vol. 140, núm. 2, p. 212-214. ↩















