Última actualizacón: 19 marzo 2018 a las 11:01
Como vimos al final de la primera parte de esta anotación, la maquinaria metabólica de los animales muestra una importante variación alométrica en relación a su tamaño: aunque la tasa metabólica aumenta de manera uniforme con la masa del organismo, este incremento no es proporcional.
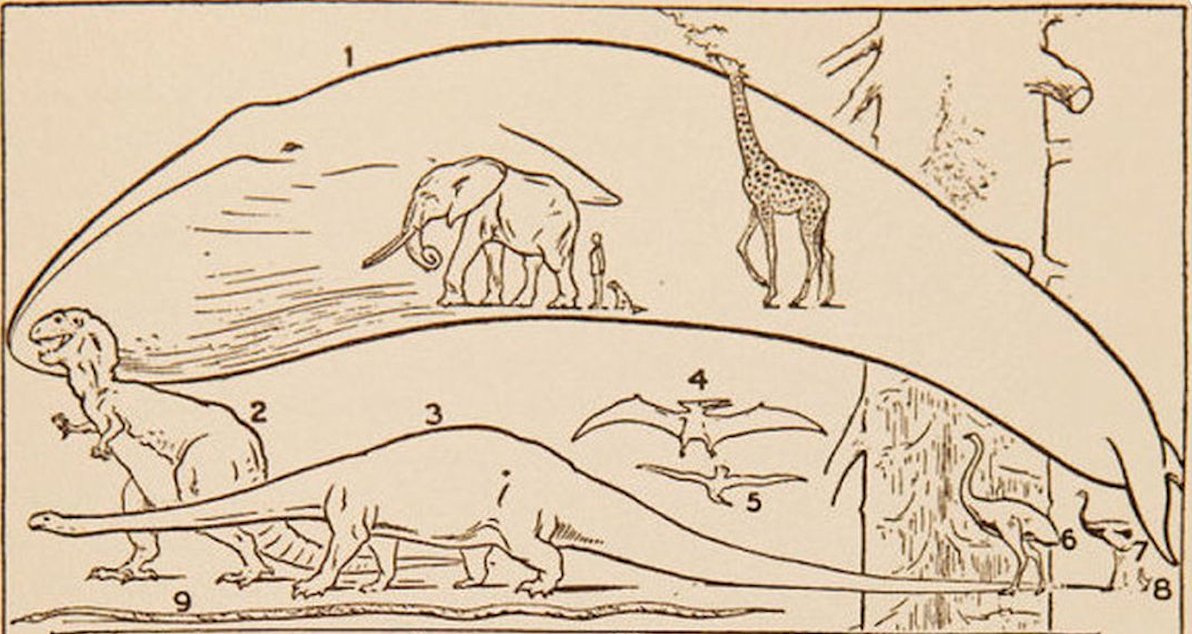
Los animales más grandes (con una mayor masa) necesitan más energía para sobrevivir que los más pequeños. Uno podría pensar que un elefante, que es unas 1000 veces más pesado que un ratón, necesitaría unas 1000 veces más energía que éste (lo que implicaría un aumento isométrico). Sin embargo, esto no es lo que sucede: cuanto mayor es el animal, aunque precise más energía, consume proporcionalmente menos que un animal de tamaño pequeño. ¿Cuál es la explicación de este fenómeno?
Los fisiólogos fueron los primeros en darse cuenta de las dificultades que entrañaba explicar el comportamiento del metabolismo en relación con el tamaño corporal. Alrededor de 1830, dos franceses, Jean-François Rameaux (fisiólogo) y Pierre Frédérique Sarrus (matemático), estudiando la temperatura de los animales de sangre caliente, llegaron a la conclusión de que la pérdida de calor debería ser proporcional a la superficie del animal (ya analizamos esto al hablar de la evaporación a través de la piel); y que tanto la ganancia como la pérdida de calor debían de ser iguales ya que el organismo mantiene una temperatura constante.
Siguiendo esta argumentación, el calor generado por el metabolismo y el perdido por evaporación deberían variar por igual con el área de la superficie (que es el cuadrado de las dimensiones lineales del animal). Sin embargo, los datos obtenidos en el laboratorio eran paradójicos ya que, si bien el calor perdido variaba con el área superficial, el generado por el metabolismo lo hacía en función del volumen corporal (el cubo de las dimensiones lineales del animal).
Otro fisiólogo, el alemán Carl Bergmann, aclaró que la clave no estaba en distinguir entre animales de sangre caliente o fría, sino entre animales de temperatura constante o variable (de hecho, Bergmann fue quien acuñó los términos homeotermo y poiquilotermo que seguimos utilizando hoy en día). Concluyó que el animal más pequeño produce más calor (por unidad de masa) que el animal grande para mantener el ritmo de la pérdida superficial, y que esta producción extra de calor significa más gasto de energía, más consumo de alimento y más trabajo realizado. Dicho de otra forma, dado que los animales homeotermos tienen que mantener la temperatura corporal constante, para compensar la pérdida de calor con el medio, deben incrementar el generado por el metabolismo. Un ser humano consume diariamente alrededor de una quinta parte de su propio peso corporal en alimento, pero un ratón debe comer la mitad de su peso al día; de ahí que su ritmo de vida sea más rápido, se reproduzca más deprisa y envejezca y muera antes.
La desventaja del tamaño pequeño es mayor cuando la pérdida de calor se ve acelerada por la conducción (como sucede por ejemplo en el Ártico) o por convección (como en el mar). De esta forma podemos comprender porqué en las grandes latitudes viven aves de gran tamaño pero no pequeñas, y también porqué viven osos pero no ratones. Es lo mismo que sucede en el mar donde no hay mamíferos de pequeño tamaño.
A pesar de estos avances, no fue hasta que entró en escena un químico agrícola suizo cuando dispusimos de análisis cuidadosos de la relación exacta entre la producción de calor (o metabolismo) y el tamaño corporal.
La ley de Kleiber

Max Kleiber se incorporó al departamento de zootecnia (una rama de la agricultura relacionada con la producción y el cuidado de animales) de la Universidad de California en Davis en 1929, y se impuso la tarea de construir unas cámaras de respiración para realizar investigaciones sobre el metabolismo de los animales. Esta estimación de las tasas metabólicas tenía importantes repercusiones en el negocio ganadero ya que permitía predecir con cierta fiabilidad la cantidad de alimento que necesitarían éstos (fundamentalmente vacas) y la cantidad de carne que producirían una vez sacrificados.
Siguiendo esta línea de investigación ―y tras innumerables mediciones con gallinas, conejos, caballos, personas etc.― llegó a la conclusión de que el metabolismo de un animal ―su respiración, su circulación sanguínea y digestión, entre otros procesos― fuera cual fuera el animal, se podía predecir tomando su peso corporal elevado a la potencia de ¾. Más aún, la tasa metabólica basal, se midiera por la tasa de consumo de alimento, consumo de oxígeno o producción de calor, variaba de un modo incluso más regular cuando la comparación no se realizaba dentro de la misma especie, sino entre especies diferentes. Así, este fenómeno de escala se reducía a un algoritmo matemático invariable, la escala de la cuarta potencia: si se pone en relación la masa y el metabolismo dentro de una escala logarítmica aparece una línea perfectamente recta que parte de ratas y palomas y va subiendo conforme aumenta el tamaño del organismo.
Veamos como lo explicó el propio Kleiber en un artículo que se ha hecho famoso, Body size and metabolism, publicado en 1932 en la revista Hilgardia (la primera publicación técnica de la Universidad de California relacionada con la agricultura):
Podemos encontrar una formulación general de la ley que expresa la relación entre el tamaño corporal y el metabolismo si representamos en un gráfico el logaritmo del metabolismo frente al logaritmo de la masa corporal. Aparece una línea recta, indicando que el logaritmo del metabolismo basal es proporcional al logaritmo de la masa corporal.
De la función linear de los logaritmos del metabolismo y la masa corporal se sigue que el metabolismo por unidad de una cierta potencia de la masa corporal es constante. Éste, de hecho, es el resultado que se obtuvo probando diferentes cálculos que se muestran en la tabla y resultó que la potencia de ¾ de la masa corporal era la unidad que encajaba mejor.

Una de las conclusiones más hermosas de esta ley según el divulgador George Johnson es que el número de latidos de una vida tiende a ser el mismo en todas las especies; lo que sucede es que los animales de mayor tamaño se toman más tiempo en agotar los suyos.

Terminaremos este apartado con una maravillosa constatación de esta ley. El fisiólogo inglés Douglas Wilkie llamó la atención, en un simposio celebrado en la Universidad de Cambridge en 1975, acerca de un hecho muy relevante en relación con la tasa metabólica de los niños inmediatamente después del nacimiento. Las mediciones mostraban que la tasa basal de consumo de oxígeno en el neonato aumentaba alrededor de 36 horas tras el parto, de 3,5 mm por minuto y kilogramo (apropiada para el tamaño de la madre) a unos 7 mm por minuto y kilogramo (apropiada para la masa corporal del bebé). Mientras el feto está dentro de la madre se comporta metabólicamente como si fuese uno más de sus órganos, acompasándose a la intensidad metabólica relativamente baja determinada por el peso de ella. En las 36 horas que siguen al nacimiento, las tasas de actividad de sus enzimas y mitocondrias han cambiado, acelerando sus procesos lo suficiente para llegar, en el segmento de la línea de Kleiber correspondiente a los humanos, al punto determinado por su propio peso como un ser pequeño y autónomo.
¿Tiene explicación?
Una explicación a los fundamentos de la ley de Kleiber podemos buscarla en la física y en la geometría del sistema circulatorio de los animales. Los animales jóvenes (y también los más pequeños) respiran más veces por unidad de peso que los más viejos (o los más grandes) de la misma especie debido al gasto energético necesario para el crecimiento; del mismo modo, los animales adultos de pequeño tamaño de una especie determinada respiran más veces por unidad de peso que un adulto de mayor tamaño de otra especie diferente debido a que una mayor parte de su masa corporal está formada por la estructura que le sirve de soporte (esqueleto, musculatura etc.) más que por reservas; la masa estructural requiere mayores costes de mantenimiento, mientras que la masa de reserva no.
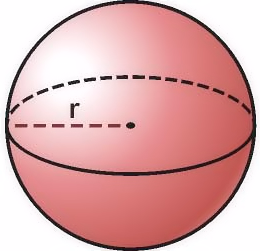
Todavía se discute por la comunidad científica si el exponente de la ley de Kleiber debe ser ⅔ en lugar de los ¾ que se ha aceptado de forma más general. Debido a que la ley de Kleiber se refiere a la obtención, uso y pérdida de energía por un sistema biológico, la tasa metabólica del sistema se tomó en principio como ⅔ (en lo que se conoce como ley de la superficie, que ya analizamos en la primera parte de esta anotación), porque se pensaba en la energía en términos de energía térmica. Los investigadores tomaron como modelo los animales esféricos para elaborar sus hipótesis ya que es una buena aproximación matemática (en términos generales podemos considerar el cuerpo de los diferentes animales como esferas de mayor o menor tamaño). Los dos tercios expresaban la relación entre el cuadrado del radio (el área de la superficie) y el cubo del radio de la esfera (que representa el volumen): cuando aumenta el radio, el volumen aumenta más rápido que el área de su superficie. Presumiblemente, este es el motivo por el que las grandes criaturas viven más tiempo que las pequeñas: a medida que se hacen más grandes, pierden menos energía por unidad de volumen en forma de calor radiado a través de la superficie.
Sin embargo, el problema con el exponente de ⅔ era que no estaba de acuerdo con muchos de los datos obtenidos empíricamente.
Forma, función y evolución de los organismos vivos
En esta nueva investigación, los autores plantean una derivación de la ley de Kleiber que opera de forma independiente a la enorme variedad de especies. Los grandes organismos multicelulares más comunes en la tierra, las plantas y los animales, presentan formas distintas según impone la distribución de la masa sobre el volumen (los animales son capaces de moverse y son aproximadamente homogéneos en su distribución masiva; mientras que las plantas son organismos enraizados con una geometría heterogénea ya que la masa del árbol está más concentrada en el tallo y en las ramas que en las hojas). Por lo tanto, la pregunta es, dadas las similitudes, ¿qué subyace al exponente dominante de ¾?
Los investigadores entienden que hay un factor que falta en la ecuación. Algunos han propuesto que tiene que ver con el espacio ocupado por los órganos internos. Otros han planteado que la fractalidad está en el fondo de la escala alométrica, una forma que es común a las ramas del árbol y los vasos sanguíneos de los animales. En este sentido, los matemáticos se dieron cuenta de que en nuestro interior existe una estructura fractal que distribuye la energía por el organismo. El sistema circulatorio, las venas, arterias y capilares son autosimilares, es decir, si examinamos el conjunto del sistema a escalas cada vez más menores, vemos que está formado por copias más pequeñas de la misma figura. Los modelos teóricos presentados por Geoffrey West, Brian Enquist, y James Brown (en su artículo A general model for the origin of allometric scaling laws in biology) pretenden mostrar porqué el exponente de ¾ puede surgir de las restricciones que impone distribuir los recursos a través de redes de ramificación jerárquica.

Ejemplos esquemáticos de los segmentos de las redes de distribución biológica: (A) los sistemas circulatorio y respiratorio de mamíferos compuestos por tubos ramificados; (B) sistema vascular de las plantas compuesto por vasos divergentes, (C) representación topológica de este tipo de redes, donde k representa el orden del nivel, comenzando con la aorta (k = 0) y terminando con los capilares (k = N), y (D) parámetros de un tubo típico en el nivel k-ésimo. Tomado de West et al. (1997).
Sin embargo, otros investigadores argumentan que, aunque las redes circulatorias pueden suponer ventajas, la potencia de ¾ tiene que ver con el transporte dirigido de nutrientes. En cualquier caso, algunos estudios recientes demuestran que puede surgir la escala de la cuarta potencia incluso cuando no hay fractalidad subyacente.
Para los autores del nuevo estudio el factor desconocido es otro: la velocidad a la cual los nutrientes son distribuidos por todo el cuerpo y se elimina el calor.
A pesar de su evolución independiente y de poseer diferentes metabolismos, las plantas vasculares y los animales bilaterales comparten importantes características de diseño: una masa interna que comprende células organizadas capaces de actividades metabólicas y bioenergéticas, un mecanismo de transporte para la distribución de las moléculas y la energía, y una superficie capaz de intercambiar esa materia y energía con el medio ambiente. Independientemente de las diferencias de forma observadas entre estos dos grupos, la física asociada a la transformación, el transporte y el intercambio de materia y energía debe imponer inevitablemente limitaciones físicas en sus diseños (un organismo es similar a un motor ―parte de la energía obtenida de la alimentación se utiliza para el funcionamiento del organismo, el crecimiento, la reproducción, mientras que el resto se disipa a través de su superficie).
Siguiendo con la notación tradicional, toman como tasa metabólica basal «B», y la masa como «M». La tasa metabólica es una medida de la energía que se distribuye por el organismo por unidad de tiempo y debe ser proporcional a la energía disipada a través de la superficie. Lo que hacen es derivar la ley de Kleiber tomando en consideración el consumo de energía, y estudian el papel de la geometría, según la relación entre la superficie y el volumen, teniendo en cuenta la energía expulsada.
Lo que han hecho los investigadores ahora es centrar su atención en esta geometría y en las limitaciones que impone a la fisiología de los organismos. Ya hemos señalado que el enlace entre los distintos organismos se encuentra en el hecho de que la tasa metabólica es proporcional al área de la superficie, y el punto crucial sobre el que llaman la atención en este estudio es que la constante de proporcionalidad tiene que incluir la velocidad de la entrega de nutrientes y/o el transporte de energía en la superficie.
Según esta aproximación, las dos cantidades que determinan la tasa metabólica «B» son el área de superficie «S» y la velocidad de transporte de los nutrientes «v» de donde obtenemos que
B ~ Sv
Cabría esperar una ley de potencias pura en la relación entre la tasa metabólica y la masa solo cuando el producto de la superficie y la velocidad de transporte guardasen proporción precisamente como una ley de potencias de la masa del organismo. Del mismo modo, sería lógico esperar que la capacidad de transporte en la superficie pudiera variar de especie a especie dependiendo tanto de las condiciones de la superficie como del ambiente. Por este motivo, un comportamiento puro de la ley de potencias únicamente podría sostenerse en una situación idealizada.
En definitiva, la relación entre el volumen y la tasa metabólica es diferente para geometrías diferentes, lo que conlleva profundas consecuencias para la vida.
Ya hemos visto que podemos caracterizar la superficie de un organismo «S» como proporcional a su volumen «V» con un exponente dado (S ~ V x). En este sentido, la naturaleza nos dice que este exponente («x») es 1 para los árboles y ⅔ para los animales. Teniendo en cuenta estos datos, los autores han elaborado la siguiente tabla:

En ella se muestran los exponentes que marcan la proporción entre el concepto reflejado en la primera columna con la masa del organismo «M». Constatamos que la variación alométrica de la tasa metabólica basal, el tiempo biológico (por ejemplo, el tiempo que dura un ciclo de circulación sanguínea o la duración de la vida), y la tasa biológica (como las tasas de mutación o las tasas de bombeo) no dependen de la geometría del organismo y son universales (vemos por ejemplo que el exponente que relaciona la tasa metabólica con la masa es idéntico —¾— tanto en árboles como en animales).
Para un árbol el volumen es toda su superficie (S ~ V) con las hojas actuando como unidades terminales donde el agua que sube desde el suelo se evapora; y cuya velocidad de transporte es constante e independiente de la masa organismo (algo imprescindible dado que el agua sube por el tronco teniendo que vencer la fuerza de la gravedad). Por otro lado, la densidad aumenta con la masa ya que, como hemos apuntado, ésta no se distribuye de manera uniforme sino que se concentra en el tronco y en las ramas. Debemos tener presente igualmente que las hojas de las plantas absorben la mayoría de la luz visible que incide sobre su superficie. Sin embargo, más allá de 700 nanómetros (justo en el límite de nuestra percepción visual), las plantas reflejan casi toda la luz solar gracias a lo cual evitan sobrecalentarse.
En contraste, un animal tiene una densidad uniforme ―la masa y el volumen son proporcionales; y la velocidad del flujo sanguíneo aumenta proporcionalmente con la masa en una tasa determinada por la masa elevada a 1/12 (M1/12). Este es el motivo por el que los animales necesitan una bomba, el corazón, para distribuir la sangre por todo el cuerpo. “Los animales necesitan ajustar el flujo de nutrientes y de calor a medida que cambia su masa para mantener la mayor eficiencia energética posible. Por eso los animales necesitan una bomba, un corazón, y los árboles, no”. Al incluir esa información en su ecuación, los expertos encontraron que habían alcanzado una explicación completa de la ley de Kleiber.

Dada la riqueza de las estrategias adaptativas de la naturaleza y las diferencias en la historia de vida, el clima, las estrategias metabólicas, y el hábitat de los organismos multicelulares, un resultado notable es que, a pesar de las desviaciones y variaciones, se observa una fuerte tendencia en la relación entre la tasa metabólica y la masa corporal. La figura de arriba muestra la gráfica logarítmica de las relaciones entre la tasa metabólica y la masa para árboles (A) y mamíferos (B). A pesar de que las curvas de las dos gráficas son distintas, en el límite mayor de la masa tanto árboles como mamíferos siguen aproximadamente la ley de Kleiber.
Consideremos ahora el otro caso limitante de la geometría del organismo que corresponde a un área de superficie mínima para un volumen dado: un animal con x = ⅔ (S ~ V ⅔). Para que la eficiencia metabólica de los animales coincida con la de los árboles, se requiere que la velocidad de transporte de los nutrientes sea proporcional a la masa del organismo elevada a 1/12 (v ~ M1/12).
Y esto es justamente lo que apoyan los datos empíricos ―una velocidad que aumenta con la masa del organismo hace precisa la existencia de un corazón. A diferencia de los árboles, el volumen de la red circulatoria de un animal no reside por completo en su superficie. Por lo tanto, se requiere una red de circulación más compleja porque la disipación sí que se produce en la superficie.
El artículo concluye con un análisis evolutivo del desarrollo de estos mecanismos. El estudio del desarrollo biológico nos lleva a la conclusión de que los patrones de simetría de los organismos, plantas y animales incluidos, se establecen durante las primeras etapas de su desarrollo. Posteriormente, estos organismos crecen y adoptan formas complejas gracias a unos mecanismos que evolucionaron de forma independiente, como la modularidad, la estructura fractal, y la segmentación. A pesar de la diversidad de formas de vida, la validez aproximada de la ley de Kleiber ofrece una notable visión unificadora de todos ellos.
Por ejemplo, en la mosca de la fruta Drosophila, un gen llamado tinman desempeña el papel de interruptor que inicia el desarrollo del corazón simple del insecto. A pesar de las importantes diferencias que presentan los sistemas circulatorios y las estructuras cardíacas de los insectos frente a los vertebrados, el gen Nkx2-5, homólogo a tinman, juega el mismo papel en el desarrollo del corazón de los vertebrados. La presencia de estos genes homólogos y las redes reguladoras asociadas en los principales linajes bilaterales hace pesar que el antepasado común desarrolló una importante innovación en el diseño, es decir, un sistema circulatorio rudimentario con una bomba cuya regulación vino dada por un gen antepasado de tinman/Nkx2-5.
Por su parte, las primeras plantas evolucionaron a partir de algas para convertirse en los primeros invasores multicelulares de la superficie terrestre. Ellas ―y sus descendientes, las modernas briofitas― carecían de un sistema de transporte complejo, es decir, un xilema conductor de agua, y un floema conductor de azúcar. De manera parecida a lo que sucedió en los animales, la evolución en las plantas de estos complejos sistemas de transporte trajo consigo un cambio dramático en su forma. En ese momento, varios linajes de plantas vasculares obtuvieron de forma independiente hojas bilaterales especializadas para la fotosíntesis y la evapotranspiración, así como raíces cilíndricas especializadas para la absorción de agua y de iones.
Este post participa en la XXX Edición del Carnaval de Biología que acoge Activa tu Neurona
![]()
Artículo principal
Banavar JR, Cooke TJ, Rinaldo A, & Maritan A (2014). Form, function, and evolution of living organisms. Proceedings of the National Academy of Sciences of the United States of America, 111 (9), 3332-7 PMID: 24550479
(Ver artículo completo aquí)
Otras referencias
Kleiber, M. (1932), «Body size and metabolism«. Hilgardia, núm. 6, p. 315-353.
West G. B., Brown J. H. y Enquist B. J. (1997) A general model for the origin of allometric scaling laws in biology. Science, vol. 276, núm. 5309, p. 122-126.