Hace tiempo que hemos superado el estereotipo que presentaba a los Neandertales como unos cavernícolas brutos y estúpidos. Las investigaciones de los últimos años han puesto de manifiesto que nuestros antepasados desarrollaron lo que se ha venido en llamar pensamiento complejo: enterraban a sus congéneres siguiendo un ritual funerario; decoraban su cuerpo con pinturas de diversos colores (rojo y negro sobre todo); utilizaban adornos en las muñecas, collares e, incluso, se ha planteado la hipótesis de que utilizaban cintas, plumas y garras de águilas con el objetivo de realzar su aspecto de la misma forma en que lo han venido haciendo los nativos americanos. Además, estudios recientes confirman que poseían las adaptaciones necesarias para el habla —aunque no se ha podido verificar aún que hubieran desarrollado un lenguaje para comunicarse—.
Tras décadas de avances en la comprensión de estos parientes en nuestro árbol evolutivo (recordemos que en 2010 se presentó el borrador de su genoma), hay varias cuestiones que todavía necesitan una respuesta: ¿cuándo se extinguieron? ¿Cuál fue el último lugar donde habitaron? Y no menos importante, ¿cuál fue la causa de su desaparición?
En los círculos académicos se vienen planteando dos teorías contrapuestas que tratan de responder estos interrogantes: la primera sostiene que los Neandertales constituían una variante arcaica de nuestra especie y que fue asimilada por las poblaciones de Homo sapiens que llegaron al continente euroasiático provenientes de África. La otra teoría defiende en cambio que los Neandertales formaban una especie genuina, y que los humanos anatómicamente modernos (AMHs por sus siglas en inglés) provocaron su desaparición al expandirse y ocupar el territorio Neandertal.
Los primeros estudios que analizaron el ADN Neandertal rechazaron la posibilidad de una hibridación entre los Neandertales y los humanos anatómicamente modernos. De esta forma ganaba peso la hipótesis de quienes afirmaban que el “choque” tecnológico y cultural entre ambos grupos fue el detonante de la extinción de los primeros. Por lo tanto, no hubo un contacto pacífico.
Por otro lado, las dataciones cada vez más precisas de los diferentes yacimientos arqueológicos permitían sostener que tras la llegada de Homo sapiens a Europa hace unos 40.000 años, los Neandertales no desaparecieron bruscamente, sino que sobrevivieron durante más de 15.000 años en distintas regiones. ¿Qué postura tomar ante estas pruebas en principio contradictoras? ¿Estamos frente a un callejón sin salida?
En realidad no todo son controversias. En lo que sí están de acuerdo la mayoría de los científicos es en que la causa final de la extinción de los Neandertales, y los factores precisos que condujeron a su desaparición pudieron variar de un lugar a otro (en algunos casos pudo ser la enfermedad, en otros la falta de alimentos o incluso la endogamia por la reducción de las poblaciones).
En cualquier caso, podemos resumir los planteamientos actuales en las dos siguientes hipótesis:
La principal causa de la desaparición de los Neandertales está relacionada con el cambio climático.
Hace alrededor de 55.000 años, el clima de Eurasia empezó a fluctuar de frío a templado y a la inversa en cuestión de unas pocas decenas de años. Durante las olas de frío, en la mayor parte del territorio habitado por los Neandertales, la expansión de la cubierta de hielo y la tundra provocó una reducción progresiva de los bosques, y por ende, de las presas potenciales que les servían de alimento. En este caso, la falta de recursos alimenticios sería la principal causa de su extinción.
No se equivocan quienes argumentan en contra de esta tesis que los Neandertales ya estaban bien adaptados a los climas fríos, aunque los defensores de la tesis del cambio climático ponen el énfasis en que con anterioridad esas fluctuaciones se daban en periodos más prolongados de tiempo. Por aquél entonces las poblaciones de Neandertales contaban con el tiempo suficiente para adaptarse a las nuevas condiciones más duras. Esta vez, sin embargo, la rapidez de los cambios en el clima hizo imposible que se recuperaran.
De esta forma, hace unos 30.000 años apenas sobrevivían en la península Ibérica pequeños núcleos aislados de poblaciones de Neandertales. En esta región había un clima más suave y era rica en recursos. En definitiva, a pesar de su capacidad de adaptación, lo que sucedió es que esos grupos eran demasiado pequeños como para asegurar su supervivencia a largo plazo.
Los Neandertales fueron superados por los Homo sapiens invasores (AMHs), que acabaron con ellos bien de forma directa, bien de forma indirecta.
Esta teoría considera que los Neandertales se extinguieron porque sucumbieron ante la superioridad, no sólo tecnológica sino cultural, de los Homo sapiens que habían emigrado de África. Sin embargo, como hemos apuntado al inicio de este artículo, cada vez hay más datos que permiten sostener que los Neandertales habían desarrollado algunos de los comportamientos que hasta entonces se atribuían en exclusiva a nuestra especie. Es erróneo considerar a los Neandertales menos “desarrollados” que aquellos.
La nueva investigación
Establecer cuál fue la relación espacio-temporal entre las poblaciones de Neandertales y AMHs es crucial para entender el proceso, el momento y las razones que condujeron a la desaparición de los Neandertales, así como conocer si fue posible algún tipo de intercambio cultural y genético entre ambas.
Si bien la meta estaba perfectamente clara, hasta ahora los desafíos técnicos habían dificultado enormemente conseguir dataciones fiables de esta época, en gran parte porque el método de datación mediante radiocarbono alcanza su límite de validez en unos 50.000 años.
Sin embargo, una nueva investigación ha aplicado técnicas de espectrometría de masas con acelerador para construir unas cronologías más exactas y fiables y así explorar el ritmo de extinción de los Neandertales (los espectrómetros de masas detectan átomos de elementos específicos —en nuestro caso carbono 14— de acuerdo a sus pesos atómicos, ignorando los abundantes isótopos que inundan esa señal —como los del nitrógeno 14 por ejemplo—).
En este sentido, el principal objetivo del trabajo publicado en la revista Nature por un amplio equipo de investigadores ha consistido en establecer una cronología y datación precisas de los yacimientos Musterienses de Eurasia ya que éstos contienen las mejores pruebas de la sustitución de un grupo humano (los Neandertales) por otro (AMHs). Debemos señalar que el Musteriense es un complejo tecnológico que incluye herramientas de piedra, hueso y otros utensilios, cuya fabricación se atribuye a los Neandertales —aunque hay algunos yacimientos cuya autoría aún se discute por los especialistas—.
Secuencias temporales
Al mismo tiempo se han analizado las llamadas “industriasde transición”, vinculadas al Paleolítico Superior aunque, en este caso, la cuestión de si fueron los AMH o los Neandertales quienes las elaboraron es una cuestión difícil de resolver (concretamente, se han analizado dos de estas industrias, la Uluzziense que aparece en Italia y Grecia, y el Chatelperroniense de la región francocantábrica).
Bien, una vez clara la metodología a emplear, los investigadores tomaron muestras de un total de 40 yacimientos arqueológicos repartidos por Europa y Eurasia occidental. El material objeto de datación fue cuidadosamente seleccionado en cada yacimiento, tomando en consideración los objetos de los estratos superiores para obtener de esta forma la datación de los más recientes en el tiempo, y por lo tanto, poder establecer el límite mínimo de antigüedad de los Neandertales que los fabricaron.
El análisis de estos datos ha confirmado que el Musteriense llegó a su fin en el rango de 41.030-39.260 años BP (Before Present, antes del presente) en toda Europa (todas las fechas del estudio están calibradas y se ofrecen con un 95,4% de probabilidad). Los datos combinados sugieren que el Musteriense terminó en un momento muy similar en todos los yacimientos desde el Mar Negro y Oriente Próximo a la costa atlántica. También se ha podido constatar que las sucesivas industrias arqueológicas “de transición”, una de las cuales se ha relacionado con los Neandertales (Chatelperroniense), llegaron a su fin en el mismo momento.
La comparación de estos datos con los resultados obtenidos del estudio de los yacimientos más antiguos de AMHs en Europa (asociados al complejo tecnológico Uluzziense) ha permitido concluir que existió un solapamiento temporal entre los dos grupos, es decir, que los humanos modernos y los Neandertales convivieron en un periodo que oscila entre los 2.600 y los 5.400 años. Esto demuestra que hubo tiempo suficiente para la transmisión de comportamientos culturales y simbólicos, así como para posibles intercambios genéticos entre los dos grupos (intercambio de ADN que ha sido confirmado por los estudios recientes de las secuencias de nuestro genoma, que sugieren que Neandertales y AMHs se cruzaron fuera de África, dando lugar a una introgresión de entre el 1,5 y el 2,1% del ADN de Neandertal en todas las poblaciones modernas no africanas).
En definitiva, la conclusión que podemos extraer del análisis global de estos datos es que en lugar de un modelo de rápida sustitución de los Neandertales europeos autóctonos por los AMHs inmigrantes, parece que el panorama fue más complejo, caracterizado por un mosaico biológico y cultural que se prolongó durante varios miles de años en diferentes regiones.
El trabajo publicado en Nature no va más allá. No desentraña las causas de la extinción de los Neandertales pero, como hemos visto, sí ofrece respuestas —aunque parciales— a las cuestiones de si hubo entrecruzamiento y si los humanos anatómicamente modernos fueron los causantes de su desaparición (en cualquier caso, recomiendo encarecidamente leer con detenimiento el material complementario que acompaña al artículo principal y que son más de 160 páginas. Sin duda, una información que no podemos dejar de lado).
En conjunto, los datos completan el marco que los estudios genéticos vienen ofreciendo en los últimos años, pero parece evidente —y los propios autores del estudio lo señalan— que es necesario realizar más excavaciones para localizar nuevos yacimientos, aplicar las técnicas de datación más modernas y así poder completar la imagen aún fragmentaria de la extinción de quienes fueron nuestros más cercanos antepasados.
Hace unos días se publicó un artículo en la revista Nature communications bajo el título The evolution of human and ape hand proportions («La evolución de la proporciones de la mano humana y de los simios»). En él, los autores han analizado las proporciones internas de la mano (la longitud de los huesos largos del pulgar y el cuarto dedo) en relación con el tamaño corporal de una muestra de primates actuales y extintos. La muestra la conforma un total de 274 primates entre los que se incluyen macacos, mandriles, gibones, orangutanes, gorilas, chimpancés y Homo sapiens; además de algunas especies extintas de homininos (Ardipithecus ramidus, Australopithecus sediba y Homo neanderthalensis).
Una de las principales características que distingue nuestras manos de las de los simios son nuestros pulgares, que son más largos en relación con el resto de los dedos. Los autores sostienen que esta simple dicotomía no proporciona un marco adecuado para probar las diferentes hipótesis acerca de la evolución humana, así como para reconstruir la morfología del último antepasado común (LCA por las siglas en inglés) de los seres humanos y los chimpancés. Y creo que esto no hacía falta ni mencionarlo, ya que cualquiera estará de acuerdo con esta afirmación: se me escapa cómo el análisis morfológico y de proporciones relativas de una parte tan concreta —aunque importante— de la anatomía humana pueda bastar para responder una cuestión de tan amplio alcance como cuál ha sido la historia evolutiva de nuestra especie.
En cualquier caso, una vez analizados los datos, los investigadores concluyen que existe una gran variabilidad en las proporciones de las manos de los hominoideos modernos, que ellos explican por diferentes procesos evolutivos: los hilobátidos muestran una mano autapomórfica debido al extremo alargamiento de los dedos y del pulgar; los chimpancés y los orangutanes por su parte exhiben una adaptación convergente relacionada con el alargamiento de los dedos (en un grado menor que los hilobátidos). Por último, los gorilas y los homininos (entre los que nos encontramos nosotros) experimentaron pocos cambios en comparación con los anteriores.
En definitiva, afirman que los resultados apoyan la idea de que la existencia de un pulgar largo en relación con el resto de los dedos que caracteriza nuestra mano (y la de los australopitecinos por ejemplo) precisó de pocos cambios desde el último antepasado común que compartimos con los chimpancés, y que esta característica fue adquirida en convergencia con otros antropoides altamente diestros como los capuchinos y los babuinos gelada.
(a) Se muestran a misma escala los dibujos de la mano de un chimpancé y humana. (b) La longitud relativa del pulgar y el cuarto dedo (menos la cuarta falange distal). La caja representa el rango intercuartil, la línea central representa la mediana, los bigotes representan el rango de valores atípicos y los puntos son los valores extremos. Tomado del artículo que se está analizando.
Dado que esa proporción es tan antigua, postulan que estos datos refutan la hipótesis de que esos cambios en la morfología de nuestra mano se produjeron como consecuencia directa de presiones de la selección natural para la fabricación de herramientas de piedra, rechazando esta visión hoy por hoy mayoritaria.
Analicemos con más detalle la anatomía de la mano para comprender mejor esa afirmación. Mientras que las manos de los primates tienen aproximadamente el mismo número de huesos, su tamaño, y por ende, sus proporciones relativas varían bastante en función de sus capacidades de locomoción. Por ejemplo, en las especies que se desplazan por los árboles, los dedos, y especialmente las falanges, son relativamente más largos que en las especies terrestres. De hecho, muchos primates arbóreos han perdido el pulgar, mientras que los lémures han visto reducido su dedo índice para permitir una mejor habilidad de agarre entre el pulgar y los dedos más laterales.
Cuando analizamos la forma de manipular los objetos, vemos que los gorilas y los chimpancés tienen dedos largos y curvos, con yemas estrechas y pulgares muy cortos. Esta disposición de la mano les permite manejar objetos de manera muy tosca, es decir, utilizan todos los dedos y la palma de la mano (para entendernos, así es como sujetamos el mango de un martillo, usando los dedos para apretarlo contra la palma de la mano), de ahí que este tipo de agarre reciba el nombre de “pinza de presión”.
Vista frontal de los huesos de los dedos de un chimpancé adulto. Atlas osteológico del chimpancé.
En cambio, nosotros tenemos un pulgar muy desarrollado con una gran movilidad y fuerza que le otorga una serie de tendones y hasta cinco paquetes musculares (nuestro metacarpiano del pulgar no sólo es más largo que el de gorilas y chimpancés, sino que también es más ancho). Del mismo modo, la segunda falange de este dedo también tiene una base muy ancha para la inserción de un músculo bien desarrollado. En cambio, tanto gorilas como chimpancés carecen de este músculo y sólo cuentan con un tendón, que realiza una función muy similar pero menos eficaz (el extremo distal de su metacarpiano del pulgar es más estrecho y en él sólo se insertan tres paquetes musculares).
Además, nuestros dedos son más cortos y terminan en unas yemas muy anchas. Una de las claves de nuestra capacidad de agarre está precisamente en las yemas, ya que al poseer abundantes terminaciones nerviosas, nos otorgan una extrema sensibilidad. Nuestro agarre recibe el nombre de “pinza de precisión”, y nos permite manipular objetos entre el pulgar y el índice de manera muy precisa. De hecho, constituye una de las bases anatómicas de nuestra habilidad para fabricar herramientas.
Representación de los músculos relacionados con el pulgar (vista superior mano izquierda Homo sapiens). 1. Aductor del pulgar. La acción de este músculo es la de aproximar el primer metacarpiano al segundo. También flexiona la primera falange sobrfe el metacarpiano. 2. Oponente del pulgar. Este músculo tira del primer metacarpiano hacia delante y adentro, permitiendo la rotación interna. Por tanto, realiza el movimiento que permite llevar el pulgar engrente de los otros dedos y efectuar las diferentes prensiones. 3. Flexor corto del pulgar. Tira del primer metacarpiano hacia dentro, hacia delante y en rotación interna. Flexiona la primera falange del pulgar. 4. Abductor corto del pulgar. Tira del metacarpiano hacia delante y dobla la primera falange sobre el metacarpiano. Composición realizada por el propio autor.
En el estudio que analizamos, los autores sostienen que los primeros homininos ya podían realizar esta pinza de precisión hace 6 millones de años, por lo que si no hemos hallado herramientas de piedra de esa antigüedad quizá sea porque nuestros antepasados no tenían las capacidades cognitivas necesarias, y no por una “incapacidad” anatómica. Bajo mi punto de vista, hacer esta afirmación tomando en consideración únicamente las proporciones de los pulgares en relación con el cuarto dedo de la mano es ir demasiado lejos. Basta examinar la complejidad de huesos, articulaciones, tendones y músculos que configuran la mano para darse cuenta que un solo rasgo no define toda la trayectoria evolutiva de una especie.
Para concluir, plantean varios escenarios evolutivos que según ellos presentan profundas implicaciones en relación con la evolución de los simios y los orígenes del ser humano:
1. Los simios existentes en la actualidad son heterogéneos en términos de las proporciones de la longitud de la mano.
Estos resultados indican una evolución paralela para el alargamiento de los dedos (donde chimpancés y orangutanes compartirían similitudes convergentes), lo que reforzaría la opinión de que las adaptaciones especializadas para la locomoción arbórea exhibidas por los simios actuales no son idénticas, es decir, que evolucionaron de forma independiente. Sostienen que este paralelismo vino facilitado por la constitución genética y desarrollo comunes de estos simios. Una de las consecuencias de esta hipótesis es que ningún simio actual serviría como análogo para describir un ancestro hipotético dado.
Esto guarda relación con las ideas previas que, junto con la proximidad filogenética entre Pan y Homo, se han utilizado habitualmente como apoyo a la hipótesis de que los homininos evolucionaron a partir de un ancestro similar a los chimpancés actuales. Las autores nos advierten con sus conclusiones que no debemos confiar en escenarios evolutivos que asuman que los simios existentes son buenos modelos ancestrales “globales”.
2. Para las proporciones en la longitud de la mano, estos resultados indican que los chimpancés y los orangutanes son convergentes; mientras que los hilobátidos evolucionaron hacia unos dedos largos de forma paralela pero con mayor intensidad, arrojando valores atípicos extremos (debido a su pequeño tamaño y a una braquiaciónespecializada).
Por lo tanto, en términos de evolución del alargamiento de los dedos, los investigadores sostienen que en algunos linajes de simios la selección natural actuó sobre la (co)variación de la longitud entre las extremidades y las proporciones de la mano en el contexto de una adaptación especializada para un desplazamiento de balanceo entre las ramas de los árboles donde se empleaban únicamente los brazos.
3. La similitud en las proporciones de la mano entre los humanos y los gorilas, teniendo en cuenta la reconstrucción de nuestro pasado africano, parece indicar que la posesión de estos dedos muy largos no fue un requisito esencial para el desarrollo del nudilleo.
4. Estas similitudes también indican que la locomoción arbórea no llegó a su fin con los australopitecinos si tenemos en cuenta la longitud de su mano.
5. Los seres humanos sólo han modificado ligeramente la longitud de sus dedos y del pulgar desde que nos separamos de nuestro ancestro común con los chimpancés hace unos 6 Ma. Esto se debió probablemente al desarrollo del bipedismo habitual en los homininos, y casi con seguridad precedió a la fabricación de las herramientas de piedra.
En definitiva, nos encontramos ante un estudio que reviste relativo interés y que ofrece unas hipótesis que requerirán de posteriores desarrollos para que puedan ser mejor perfiladas.
No quería terminar este análisis sin recomendar encarecidamente para aquellos que estén interesados en la evolución de la morfología de la mano, la lectura de un estudio publicado en la revista Journal of Anatomy el año 2008 (puedes leerlo aquí). En él, Tocheri y colaboradores sostienen que las pruebas moleculares y fósiles tienen importantes consecuencias para la interpretación de la historia evolutiva de la mano dentro de la tribu Hominini. En primer lugar, la parsimonia apoya la hipótesis de que la mano del último ancestro común de los humanos y chimpancés es más probable que es pareciera a la de un gran simio actual en general (Pan, Gorilla, y Pongo), que a la de un simio africano en particular. En segundo lugar, proporciona un contexto para la interpretación de los cambios derivados de la mano que se han desarrollado en diferentes homínidos. Sin embargo, la mayoría de las características primitivas que es probable que estuvieran presentes en el último ancestro común de Pan y Homo se mantienen en las manos de Australopithecus, Paranthropus/primeros humanos, y Homo floresiensis.
Referencias
Almecija, S.; Smaers, J. B. y Jungers, W. L. (2015), «The evolution of human and ape hand proportions». Nature communications, vol. 6, p. 1-11.
Tocheri, M. W., et al. (2008), «The evolutionary history of the hominin hand since the last common ancestor of «Pan» and «Homo«. Journal of Anatomy, vol. 212, núm. 4, p. 544-562.
La museología, como la ciencia en general, avanza a grandes pasos de la mano de la tecnología. El desarrollo imparable de internet y el enorme potencial que la informática y el procesado de la información ofrecen, están permitiendo una nueva forma de acercarnos a los muesos, lejos ya la clásica imagen de un lugar donde se amontonan objetos sin conexión entre sí. Se abre una ventana que nos permite un acceso privilegiado a los restos del pasado, en cualquier momento y en cualquier lugar.
Felipe V fundó en 1716 la Real Biblioteca o Librería Pública con un pequeño museo, un Gabinete de Antigüedades que, pasado el tiempo, se convertiría en uno de los núcleos fundacionales del Museo Arqueológico Nacional (por cierto, recientemente remodelado y modernizado). Una Orden del rey, puesta en vigor en 1713, dictaba las normas para recoger y surtir de curiosidades de historia natural a la recién creada Librería Pública:
Las antigüedades dan luz de lo que fueron los países en los tiempos más remotos y por ellas se saca el conocimiento del aumento y disminución que han tenido: con este motivo se procura investigar lo conducente a su averigüación, dando noticia de los vestigios que permanezcan en algunos parajes.
Mucho tiempo ha pasado desde entonces pero aún perdura la misma idea: es necesario preservar y estudiar los vestigios del pasado para saber quiénes somos, para conocer nuestro devenir por la historia.
En este sentido, el Museo de Historia Natural de Idaho se ha embarcado en la compleja misión de acercar y hacer totalmente accesibles sus colecciones al gran público. Para ello han creado una mediateca virtual de objetos arqueológicos y restos fósiles de muy diverso tipo, que han sido escaneados utilizando la más moderna tecnología.
El Laboratorio de Virtualización es una unidad de investigación del Museo de Historia Natural que pertenece al campus de la Universidad Estatal de Idaho. La labor de sus integrantes es documentar cada pieza y obtener una representación digital de sorprendente calidad (las imágenes pueden verse en 360o y ampliarse con una resolución superior a la que puede percibir el ojo humano). Se han propuesto escanear las colecciones del museo, los materiales procedentes de excavaciones arqueológicas y paleontológicas, restos de fauna etc.
Cráneo de coyote.
Los investigadores son conscientes de que muchos de estos objetos son perecederos o muy frágiles debido al daño que sufren por las manipulaciones repetidas. El objetivo por tanto es la creación de unos archivos virtuales de esas colecciones para preservar la integridad de las muestras y proporcionar un acceso global (y gratuito) a las colecciones.
Unido a lo anterior (y no menos importante) el laboratorio permite la aplicación de esta tecnología a la investigación, la docencia y otros proyectos desarrollados por científicos y educadores, permitiendo proyectos de colaboración interdisciplinares. Les dejo con unos vídeos que muestran el proceso de digitalización:
Ya se han escaneado más de 15.000 piezas y el trabajo continúa.
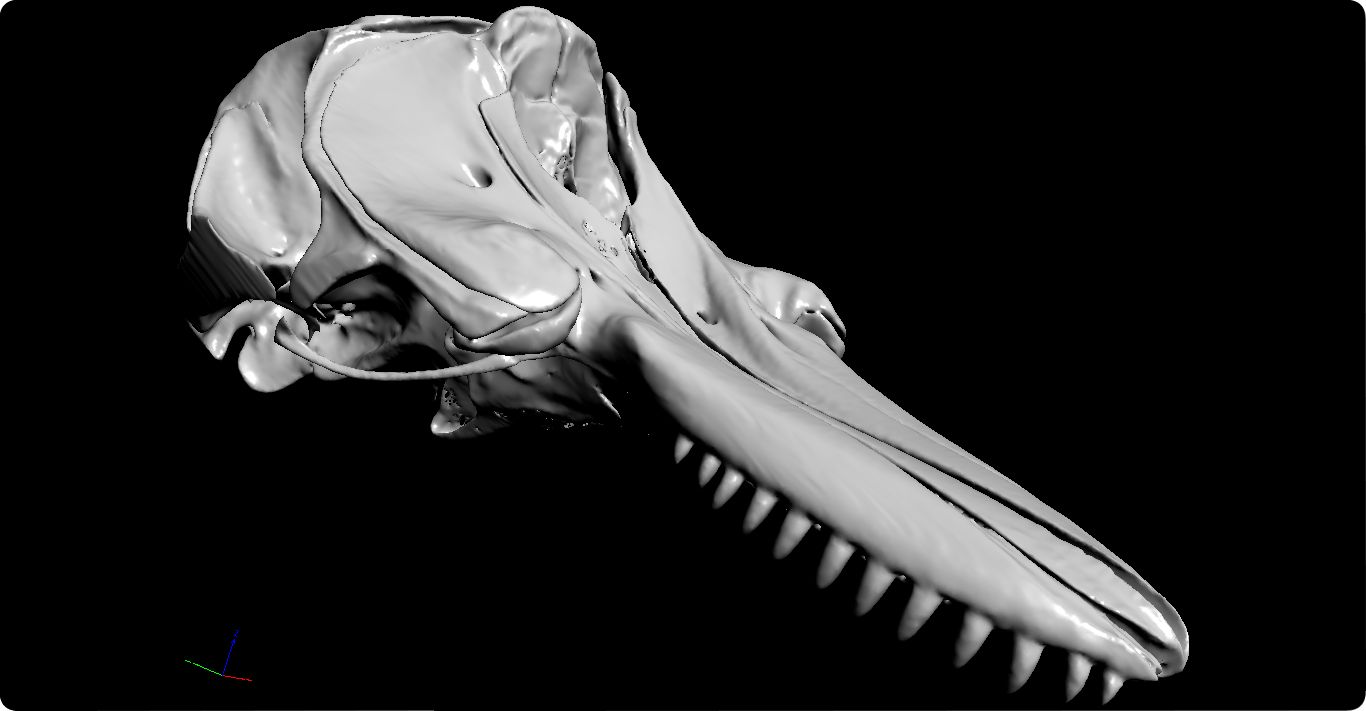
PS: Les recomiendo que curioseen por las bibliotecas digitales porque, estoy seguro, encontraran muchas curiosidades que no podían imaginar. Es lo que me ha pasado a mí cuando he visto el cráneo de la orca que aparece más arriba ya que hace unos días salió el último capítulo del podcast Catástrofe Ultravioleta que gira precisamente en torno a estos espectaculares odontocetos (que no ballenas). No se lo pierdan…
El frío viento nocturno se colaba por las rendijas de la desvencijada casa. Ese silbido agudo y el sordo entrechocar de las ventanas impedía que nadie pudiera conciliar el sueño, aunque tampoco ayudaba el hecho de que hubiese comenzado a llover con fuerza hacía apenas unos minutos. Así las cosas, los ocupantes de la casa se dedicaron a asegurar bien puertas y ventanas, así como a colocar cazos y otros cacharros allí donde el agua comenzaba a colarse por las goteras. Estaba claro que la casa había visto tiempos mejores. Cuando terminó de ayudar a su madre y a su hermano para evitar que la tormenta se llevara la casa con ellos dentro, la pequeña Mary ―con doce años recién cumplidos― se arrebujó en la cama bajo una colección de mantas de varios tamaños y colores para intentar desprenderse del pegajoso frío del cuerpo.
Pero en realidad no podía dormir, o deberíamos decir que no quería dormir. Estaba ansiosa y exultante porque sabía que esa fuerte lluvia sería su mejor aliada, una ayuda inestimable en la pesada y, en ocasiones, tediosa tarea que le esperaba al amanecer. A pesar de todo, no cambiaría su vida por nada del mundo ya que amaba el trabajo que venía haciendo desde hacía muchos años junto a su padre y su hermano mayor. Ocasiones como esta, en las que los elementos se conjuraban para verter sobre la tierra cantidades inimaginables de agua, eran en realidad las más idóneas para salir a pasear por la costa, martillo en mano, y tratar de devolver a la luz los testigos de un pasado remoto, los restos de los seres vivos que mucho tiempo atrás señorearon en ese mismo lugar y que se habían convertido en roca con el paso de los eones. Estaba convencida de que el día traería magníficas oportunidades, y con esa idea en mente, y la sensación de que su padre seguía junto a ella ―había fallecido dos años antes―, Mary cayó finalmente en un profundo sueño mecida por el rumor de las olas.
Y llegó el día. Llegó la mañana pero ningún rayo de luz se coló por las ventanas, y el cielo permanecía tan encapotado que se hacía difícil distinguir la línea del horizonte, dónde terminaba el mar y comenzaba el cielo. La grisácea monotonía sólo se quebraba por el sonido familiar del romper de las olas en la playa y en el acantilado.
―¡Joseph, rápido!, tenemos que salir cuanto antes para que nadie se nos adelante ―gritó Mary saltando de la cama y buscando ropa abrigada que ponerse― ¡Hoy va a ser un gran día!
―¿A qué tanta prisa? No ha dejado de llover en toda la noche y seguro que con este mal tiempo pocos serán los que quieran venir al pueblo ―protestó su hermano mientras se tapaba aún más con una gruesa manta― Tus piedras seguirán ahí mañana y pasado mañana y al día siguiente…
En realidad Mary no terminó de oír lo que su hermano pretendía decirle porque ya había salido de la habitación y bajaba rápidamente las escaleras al tiempo que llamaba a su perrito Tray para que saliera fuera con ella.
Lyme Regis, el pueblo donde vivían los Anning, había pasado por mejores momentos. En otro tiempo uno de los principales puertos de Inglaterra, había sufrido enormemente con el embargo comercial impuesto por las guerras que se libraban tanto en el continente contra Napoleón como en las colonias de ultramar contra Estados Unidos. En cualquier caso, Lyme había sabido sobreponerse al convertirse en un destino turístico que era visitado por ilustres personajes como la mismísima Jane Austen, quien plasmó el ambiente bucólico de la región en su novela Persuasión.
Pese a que había dejado de llover hacía poco, aún se palpaba y olía la humedad y el frío de la noche. El sol no conseguía despuntar a través de la gruesa capa de nubes pero a Mary no le afectaba, al contrario, eran el mejor presagio y lo sabía. Los acantilados se prolongaban durante muchos kilómetros a lo largo de la costa, una pared casi ininterrumpida de unos diez metros de alto que exponía claramente la disposición horizontal de los diferentes estratos, sucesivas capas de tierra y roca de diferentes grosores que mostraban, para quien sabía leer sus líneas, un detallado registro geológico así como el enorme lapso de tiempo comprimido en ellas. Estos acantilados habían brindado gran cantidad de fósiles que los hermanos Anning venían recogiendo desde hacía años junto a su padre. Una vez limpios los exhibían en un tenderete improvisado junto a la parada del carruaje que, cada cierto tiempo, dejaba grupos de turistas en la posada local, con la esperanza de venderlos a buen precio. Desde la muerte de su padre hacía dos años, los únicos ingresos de la familia provenían de la venta de estas curiosidades geológicas que no hacían sino atraer más curiosos a la costa.
Al poco rato de que Mary saliera de la casa, Joseph le dio alcance y se unió a ella y a Tray en la búsqueda de nuevos tesoros.
―Algo tiene que cambiar hermanita ―murmuró Joseph sin mirar a su hermana mientras caminaba y golpeaba el suelo con su bastón con punta de cobre.
―¿A qué te refieres?
―Estoy cansado de perder el tiempo recogiendo piedras para que nos den unos pocos peniques por ellas. Tengo tanto frío en los huesos que no recuerdo la última vez que pasé calor ―mientras se quejaba caminaba dando fuertes pisadas en un intento inútil de calentarse los pies.
Mary se detuvo en seco y miró fijamente a su hermano a los ojos.
―No te comprendo. Lo que hacemos no es ninguna pérdida de tiempo. ¿Es que ya te has olvidado de los buenos momentos que pasamos junto a padre recorriendo estas costas, todo lo que nos enseñó? Además ―añadió con un brillo repentino en la mirada― ¿no te das cuenta de que tenemos la oportunidad de rescatar del olvido el pasado? Yo tampoco quiero pasar el resto de mi vida vendiendo baratijas a los turistas, pero creo que si recuperamos y estudiamos estas “piedras” como tú las llamas, será como traer de nuevo a la vida los animales que vivieron hace muchísimo tiempo.
Mary no comprendía cómo era posible que alguien no sintiese curiosidad y fascinación por las maravillas que aguardaban en las rocas. A pesar de que no poseía una educación convencional (había aprendido a leer los domingos que asistía con sus padres a la iglesia congregacional) sabía que los fósiles eran vestigios de un pasado remoto, los restos olvidados por los hombres de antiguos seres que solo esperaban una nueva oportunidad de ver la luz. Y su tarea era precisamente esa, recuperarlos para estudiarlos y mostrarlos de nuevo al mundo.
Dibujo del cráneo de ictiosauro hallado por Joseph Anning en 1810.
―Yo lo único que digo ―continuó Joseph con su argumentación― es que en el telar necesitan ayuda. Me gustaría aprender un oficio donde trabajar sin pasarme el día tirado en el suelo revolviendo la tierra y calado hasta los huesos. Esta moda de los fósiles pasará y entonces no tendremos forma de llevar comida a casa.
Mary no prestó atención a sus últimas palabras, ensimismada como iba, y corrió adelantándose a su hermano. Habían llegado a un saliente de la costa donde se había producido un enorme desprendimiento de tierra. Una parte de la pared del acantilado se había desplomado sobre la playa, sin duda por la acción del torrente de agua caído durante la noche. Tray comenzó a trepar entre las rocas y Mary le siguió con cautela para no provocar un nuevo corrimiento de tierra. Cuando llegó a la cima y escuchó los ladridos de Tray apenas pudo contener un grito.
―¿Qué sucede?, ¿te has hecho daño? ―preguntó resoplando Joseph cuando finalmente pudo alcanzarla.
―Mira eso, te dije que hoy iba a ser un gran día.
Mary señalaba la parte baja del acantilado que quedaba al otro lado del desprendimiento. Parcialmente cubiertos de barro, y comprimidos entre varias rocas, asomaban unos enormes huesos fosilizados. De un color más oscuro que la roca circundante resaltaban con claridad y, a pesar de que todavía estaban enterrados en parte, se podían atisbar unas grandes vértebras que terminaban en lo que parecía ser la cola de un animal enorme. Con la emoción a flor de piel, los hermanos bajaron rápidamente mientras miraban extasiados el gran tamaño del esqueleto que habían descubierto.
―¡Es sorprendente! ―exclamó Mary con gran excitación― Tenemos que darnos prisa para terminar de desenterrarlo y vamos a necesitar mucha ayuda ―continuó mientras comenzaba a golpear los extremos del enorme bloque de piedra con su martillo.
―Iré a la cantera para que vengan a echarnos una mano ―apuntó Joseph mientras volvía a subir por donde habían venido― No podremos moverlo de aquí hasta que lo hayamos sacado por completo.
Ya sola y con tiempo para observar detenidamente, Mary se dio cuenta enseguida de que habían topado con algo muy importante. Golpeaba con el martillo con una delicadeza que sin duda sorprendería a cualquiera que no conociera el oficio, esa herramienta que le había fabricado su padre y sin la que no salía de casa. Viéndola ahí, en el suelo, apoyada con un brazo mientras con el otro manejaba el martillo uno se daba cuenta de que era muy buena en su trabajo, muy meticulosa y paciente, ya que sabía que un golpe en un lugar inadecuado o con más fuerza de la debida podía romper en pedazos aquellos testigos mudos atrapados en la piedra.
Esqueleto de ictiosauro.
Después de muchas horas de duro trabajo, con la ropa totalmente empapada pero con una amplia sonrisa en el rostro, Mary pudo contemplar el hallazgo en su integridad. Ante sí tenía un esqueleto de más de cinco metros de largo, con dos grandes aletas en la parte delantera y otras más pequeñas donde comenzaba la cola. Por desgracia no conservaba la cabeza pero pensó rápidamente que el cráneo que su hermano había encontrado el año anterior encajaba perfectamente con este ejemplar, tanto por el tamaño como la ubicación. ¡Qué gran descubrimiento!
Sin perder más tiempo tomó el papel y el lápiz que siempre llevaba consigo y comenzó a dibujar los restos con la mayor fidelidad posible. Mientras lo hacía no dejada de preguntarse cómo habría vivido este animal y cómo se habría alimentado para mantener un cuerpo tan enorme. Estaba claro por las aletas que era un animal marino, pero el cráneo que encontraron el año anterior mostraba unos dientes puntiagudos que sobresalían de la boca como los cocodrilos. ¿Podría respirar bajo el agua?, ¿caminaría sobre la tierra? Estas y muchas más preguntas no cesaron de abordarla, sabiendo con la certeza que sólo puede sentir alguien realmente apasionado por su trabajo, que pasaría el resto de su vida tratando de resolver todas esas incógnitas.
Post scriptum:
Mary Anning (1799-1847) tenía doce años cuando encontró el esqueleto casi completo de un ictiosaurio (cuyo cráneo descubrió su hermano Joseph un año antes), dedicando los siguientes treinta y cinco años de su vida desenterrando y analizando los fósiles de la región cercana a su casa de Lyme Regis.
Debido a su condición de mujer tuvo vetado el acceso a las instituciones científicas de la época, a pesar de que muchos artículos leídos ante la Sociedad Geológica describían sus trabajos pero sin mencionarla. Este injusto aislamiento académico no le desanimó a continuar con su labor investigadora, dibujando y diseccionando peces y sepias para comprender mejor la anatomía de los fósiles con los que trabajaba. Charles Lyell fue de los pocos científicos de la época (junto con William Buckland, Louis Agassiz y Roderick Murchinson) que reconoció y alabó su obra.
Joseph Anning, que pasó muchos años junto a su padre y su hermana buscando fósiles, se convirtió en tapicero en 1825, se casó y tuvo tres hijos.
Nota:
Todas las imágenes han sido tomadas de Wikimedia commons.
Esta entrada participa en el I Certamen de cuentos de ciencia organizado por el blog Cuantos y Cuerdas
Les propongo que hagamos un breve y sencillo experimento. Prueben a introducir en el buscador de imágenes de Google los términos “evolución”, “evolución humana” o “evolución del hombre”. Verán desfilar ante sus ojos ―en múltiples variantes pero con un claro denominador común― una de las imágenes que más daño ha hecho a la comprensión de la evolución de nuestra especie:
Sigan leyendo esta anotación porque espero que al final comprendan bien el porqué.
En la primera parte de esta serie analizamos qué es la paleoantropología y cómo se forma la “materia prima” ―por decirlo de alguna forma― de su objeto de estudio: los restos fósiles de los organismos que vivieron en el pasado. Cuando estos fósiles comenzaron a ver la luz y nació la paleontología moderna, gracias sobre todo al trabajo de Georges Cuvier, se inició un estudio sistemático de los mismos que llevó a la comprensión de que la vida tiene una historia, que existe variabilidad, y que se puede analizar el cambio en la morfología de los animales extintos. Este hecho fundamental, junto con algunos otros, llevaron al asentamiento de la idea de la evolución de las especies.
No vamos a hacer un estudio pormenorizado de las diversas corrientes que llevaron al planteamiento de la teoría de la evolución por Charles Darwin (y Alfred Russel Wallace) en 1859, ni tampoco vamos a detallar los avatares por los que ha atravesado la disciplina desde entonces ya que supondría alargar en exceso este texto. Lo que sí pretendo es ofrecer una visión clara y comprensible acerca de qué es la evolución biológica y cómo funciona.
En latín clásico, la palabra evolutio significa “desenrollar o abrir un libro” (detalle), por lo que podemos traducirla como «leer», ya que los libros y documentos eran, en época romana, rollos de papiro o pergamino que había que desenrollar para poder leerlos. El verbo relacionado, evolvere, tiene como raíz el verbo volvo que significa “girar, dar vueltas” (detalle). Lo que estos términos sugieren es una idea de movimiento, de un camino que se abre ante nosotros y que podemos seguir. Curiosamente, a pesar de que en la actualidad asociamos de forma casi inmediata el término «evolución» con la obra cumbre de Charles Darwin titulada “El origen de las especies”, éste prefería hablar de “teoría de la descendencia con modificación”; puntualizando que esa modificación no estaba predeterminada, orientada o dirigida, sino que era producida por lo que llamó la «selección natural». Tanto es así que en la primera edición de ese trascendental libro, la palabra «evolución» sólo aparece una vez, y es precisamente al final del mismo (de hecho es la última palabra de la obra):
Hay grandeza en esta concepción de que la vida, con sus diferentes facultades, ha sido alentada en unas cuantas formas o en una sola, y que, mientras este planeta ha ido girando según la constante ley de la gravitación, se han desarrollado [evolved] y se están desarrollando, a partir de un comienzo tan sencillo, infinidad de formas cada vez más bellas y maravillosas.
[…] from so a simple a beginning endless forms most beautiful and most wonderful have been, and are being, evolved.
La evolución no busca, pero encuentra *
Siguiendo los postulados de Francisco Ayala, podemos decir que la teoría de la evolución se ocupa de tres materias diferentes. La primera es el hecho mismo de la evolución, es decir, el hecho de que las especies cambian con el paso del tiempo y que es posible describir su historia. Si lo hacemos, descubrimos que todas ellas están emparentadas entre sí al tener un antepasado común. Podemos resumir este primer punto con la afirmación de que todos los seres vivos descienden de antepasados comunes, que son cada vez más diferentes de sus descendientes cuanto más tiempo ha pasado entre unos y otros.
Representación gráfica de la ascendencia común (por el autor).
La segunda materia es la historia de la evolución. A estudiar la evolución tratamos de establecer las relaciones de parentesco entre unos organismos y otros, así como determinar, de forma tan precisa como sea posible, en qué momento se produjo la separación entre los distintos linajes que llevan a las especies vivas en la actualidad (o saber en qué momento quedaron extintas). La evolución biológica implica por tanto el origen de nuevas especies.
La tercera y última materia se refiere a las causas de la evolución. Quizás sea este el punto que en la actualidad genera más debate en la comunidad científica, y también el más importante. Una vez sentado que los organismos cambian y que se diferencian cada vez más con el paso del tiempo, se vuelve esencial comprender los mecanismos o los procesos que causan esa evolución. Darwin, por ejemplo, describió el mecanismo de la selección natural, el proceso que explica la adaptación de los organismos a su ambiente y la evolución de órganos y funciones. Otros procesos evolutivos importantes son los relacionados con la genética (desconocida para Darwin).
Bueno, hasta ahora hemos hablado de muchas cosas pero no hemos explicado qué dice la teoría de la evolución. Como hemos apuntado, la evolución biológica es el proceso de cambio y diversificación de los organismos en el tiempo (y en términos genéticos, el proceso de cambio en la constitución genética de los organismos a través del tiempo). Actualmente, la teoría de la evolución combina las propuestas de Charles Darwin y Alfred Russel Wallacecon las leyes de la herencia descritas por Gregor Mendel, así como otros avances posteriores en genética. Según esta teoría, que recibe el nombre de “síntesis moderna” o “teoría sintética”, la evolución se define como un cambio en la frecuencia de los alelos de una población a lo largo de las generaciones. Este cambio puede darse por diferentes mecanismos tales como la selección natural, la deriva genética, la mutación y la migración o flujo genético (más bajo explicaremos brevemente estos conceptos).
El argumento central de Darwin parte de la existencia de variaciones que se heredan. Las variaciones favorables, desde el punto de vista del organismo, son las que incrementan su posibilidad de sobrevivir, pero no vivir por vivir, sino vivir para dejar descendencia, para transmitir su carga genética. La selección natural surge como consecuencia de diferencias en la supervivencia, la fertilidad, el ritmo de desarrollo, el éxito en encontrar pareja, o en cualquier otro aspecto del ciclo vital del organismo. Todas estas diferencias pueden ser aglutinadas bajo el término de reproducción diferencial puesto que todas ellas afectan al número de descendientes que dejan los organismos. Las variaciones favorables serán entonces conservadas y multiplicadas de generación en generación a expensas de las menos ventajosas, precisamente porque quienes las portan viven lo suficiente como para transmitirlas a su descendencia con más eficacia. A medida que las variaciones más útiles van apareciendo, irán reemplazando a las menos ventajosas, y como consecuencia de ello, los organismos cambiarán en su apariencia y configuración. Este proceso de multiplicación de variaciones ventajosas, a costa de las que no lo son, es lo que Darwin llama selección natural.
Como resultado de tal proceso, los organismos estarán mejor adaptados a su ambiente. La razón de ello es que son precisamente las variaciones que mejoran la adaptación al ambiente las que aumentan la probabilidad de que un organismo sobreviva y se multiplique. El proceso se facilita por el hecho de que los ambientes en que los organismos viven también están en continuo cambio. Estos cambios ambientales incluyen no solo los relacionados con el clima y otros aspectos físicos, sino también el entorno biológico: los depredadores, los parásitos y los competidores con los que un organismo interactúa le afectan de manera importante. Al mismo tiempo, el entorno biológico de una especie cambia sin cesar puesto que las especies que constituyen ese entorno están evolucionando a su vez. De esta forma, el proceso de evolución por selección natural implica una retroalimentación que hace que la evolución continúe indefinidamente.
Cuando Darwin publicó “El origen de las especies” no tenía ninguna prueba de la existencia de la selección natural y elaboró su teoría sólo por inferencia. Ernst Mayr ha esquematizado los argumentos de Darwin en cinco hechos y tres deducciones que son muy útiles para comprender la esencia de esta cuestión:
Hecho número uno: Todas las especies tienen una gran fecundidad potencial que debería producir un crecimiento muy rápido de las poblaciones, siempre que todos sus miembros sobreviviesen y se reprodujesen.
Hecho número dos: salvo fluctuaciones anuales u ocasionales, el número de miembros de las poblaciones se mantienen estables.
Hecho número tres: los recursos naturales son limitados y en un entorno relativamente estable se mantienen más o menos constantes (debemos señalar que por recursos se entiende tanto el espacio físico como el alimento disponible).
Primera deducción: teóricamente, el número de individuos debería sobrepasar rápidamente las capacidades de mantenimiento del entorno; este desequilibro va a producir una “lucha por la existencia” que sólo permitirá que se reproduzca una pequeña parte de los miembros de las poblaciones. En realidad, Darwin empleó el término “lucha” de forma metafísica ―a pesar de algunas interpretaciones malintencionadas― por lo que hoy hablamos de “competencia”.
Hecho número cuatro: dos individuos de una población nunca son exactamente iguales y toda población manifiesta una gran variabilidad. Esta fórmula pone de manifiesto algunos datos esenciales de la revolución darwiniana. Los autores previos manejaban un concepto de especie según el cual existe un tipo, un modelo de cada especie, que se reproduce de forma inmutable (corriente que ha venido a llamarse fijismo), donde las variaciones observadas son meros accidentes de “copia”. En cambio, para Darwin la variabilidad es una propiedad fundamental de la especie; no puede existir un tipo privilegiado puesto que todos los individuos son igualmente representativos de la especie con una constitución única.
Hecho número cinco: gran parte de esta variabilidad es heredable. La variabilidad por tanto es el material del cambio evolutivo.
Segunda deducción: la supervivencia en este ambiente de competencia no se debe al azar, sino que depende en gran parte de la constitución genética de los supervivientes. Esta supervivencia desigual constituye el proceso de selección natural (hoy sabemos que esta afirmación debe ser matizada ya que, dependiendo de las especies, la parte debida al azar o al determinismo genético en la supervivencia varía en límites muy amplios, aunque siempre existe una combinación entre ambos).
Tercera deducción: a lo largo de las generaciones, el proceso de selección natural conduce a un cambio gradual de las poblaciones, es decir, a la evolución con producción de nuevas especies. Este proceso de diversificación está contrarrestado por la extinción de las especies: se estima que casi el 100% de todas las especies que han existido en el pasado han desaparecido sin dejar descendientes. Las especies actuales, estimadas en unos 10 millones (las descritas por los biólogos son menos de 2 millones) son el saldo entre la diversificación y la extinción.
Vamos a explicar cómo opera la selección natural con un ejemplo que se ha convertido en clásico: el melanismo industrial. Tenemos una especie de polilla cuyos miembros pueden ser de color oscuro o claro, en este caso nos referimos concretamente a la polilla del abedul Biston betularia —en la imagen de abajo, la forma clara (definida como «común») está rotulada con el número 2; y la forma oscura (definida como «carbonaria») rotulada con el número 1.
En 1848, en pleno apogeo de la revolución industrial en Inglaterra, los naturalistas se dieron cuenta de que en áreas industriales, contrariamente a lo que pasaba en zonas no contaminadas, las formas oscuras de esta polilla predominaban sobre las claras.
La explicación tiene que ver con la contaminación ambiental: el hollín de las fábricas mata los líquenes grisáceos-claros que se encuentran sobre la corteza de los abedules, donde estas polillas pasan gran parte de su tiempo, haciendo al mismo tiempo que esa corteza se vuelva más oscura. A partir de esta evidencia, H. B. D. Kettlewell llevó a cabo una serie de experimentos que demostraron que las formas oscuras se camuflaban mejor de las aves depredadoras que la claras en la corteza de los árboles, siendo de esta forma favorecidas por la selección —en la imagen superior vemos una polilla común sobre una corteza de abedul sin contaminar (3), y en la imagen siguiente (4) ambas polillas sobre la corteza ennegrecida por el hollín de las fábricas. En este caso concreto, el color oscuro se convierte en un rasgo adaptativo porque sus portadores sobreviven más que los de color claro.
Pero esta ventaja de la forma oscura no tendría ninguna trascendencia si no fuese hereditaria. Dado que el color oscuro esta determinado por un tipo (o alelo) de un gen, mientras que el color claro lo está por otro alelo del mismo gen, el que las polillas oscuras dejen más descendientes implica que el alelo oscuro aumentará su representación en la población de la siguiente generación. Las adaptaciones son, por tanto, aquellas características que aumentan su frecuencia en la población por su efecto directo sobre la supervivencia o el número de descendientes que dejan aquellos individuos que la portan. En nuestro ejemplo, las formas claras eran miméticas en las zonas no contaminadas, siendo aquí seleccionadas a favor y, por tanto, más numerosas.
En este punto no quiero dejar pasar la oportunidad de traer a colación al genial Stephen Jay Gould, un gran científico y enorme divulgador, que era capaz de unir ciencia y arte cada vez que nos explicaba cualquier concepto:
la formulación básica ―el esqueleto― de la selección natural es un argumento de una simplicidad desarmante, basado en tres hechos innegables (sobreproducción de descendencia, variación y heredabilidad) y una inferencia silogística: la selección natural, o la afirmación de que los organismos con más éxito reproductivo serán, en promedio, las variantes que, por azar, resulten estar mejor adaptadas a los entornos locales cambiantes, las cuales, por herencia, transferirán a sus descendientes sus rasgos favorecidos.
Para mostrarnos gráficamente su visión de la teoría de la evolución escogió una figura procedente de la edición latina de 1747 de una obra precursora en la historia de la paleontología: el tratado de Agostino Scilla titulado La vana speculazione disingannata dal senso(La vana especulación desengañada por los sentidos). La obra fue publicada en italiano en 1670 y, entre otras bellas ilustraciones, cuenta con una etiquetada “coral articulado presente en gran abundancia en los acantilados y colinas de Messina”:
Gould nos explica que el tronco central del coral ―la teoría de la selección natural (1)― no puede cortarse porque la criatura moriría. Este tronco central se divide luego en un número limitado de ramas principales, que él llamó “los puntales básicos” (2, 3 y 4) y que se corresponden con las tres partes esenciales de la teoría darwiniana (sobreproducción de descendencia, variación y heredabilidad): son tan indispensables que la eliminación de cualquiera de ellas supondría poner en entredicho la teoría al completo de forma que sería necesario rebautizar la nueva estructura básica. Por último, hay otros posibles puntos de corte que, si bien obligan a repensar algunos conceptos, no suponen poner en peligro el esquema esencial. En estas partes más alejadas del tronco central podemos incluir el equilibrio puntuado, la teoría neutralista etc.
Mecanismos evolutivos
Ya hemos visto el principal mecanismo de cambio evolutivo, la selección natural. Pero como también hemos apuntado, existen otros que vamos a comentar brevemente a continuación (en futuras anotaciones los veremos con más detalle).
Las frecuencias génicas pueden cambiar por razones puramente aleatorias (lo que se llama deriva genética). En cada generación se produce un sorteo de genes (llamado recombinación) durante la transmisión de gametos de los padres a los hijos. La mayoría de los organismos son diploides, es decir, tienen dos copias de cada gen (una heredada de la madre y otra del padre). Sin embargo, los gametos de estos organismos portan solo uno de los dos ejemplares (alelos) de cada gen. El que un gameto lleve un alelo u otro es una cuestión de azar. Como el único componente que se transmite de generación en generación es el material genético (los genes), el que un individuo deje más descendientes implica que sus variantes génicas (alelos) estarán más representadas en la siguiente generación.
La deriva genética produce un cambio aleatorio en la frecuencia de los alelos de una generación a otra, resultando que estadísticamente se produce una pérdida de los alelos menos frecuentes y una fijación de los más frecuentes. Como es de esperar, este mecanismo provoca una disminución en la diversidad genética de la población y tiende a homogeneizar el acervo genético, motivo por el que la magnitud de los cambios está inversamente relacionada con el tamaño de la población: cuanto mayor sea el número de individuos en una población, menor será el efecto de la deriva genética. Podemos ilustrar este mecanismo con el clásico ejemplo de lo que sucede cuando lanzamos una moneda al aire. Si lanzamos una moneda dos veces, no nos sorprende que salgan dos caras, aun cuando eso significa que sale cara el 100% de las veces. Sin embargo, sí resultaría sorprendente y sospechoso que salieran 20 caras seguidas en 20 tiradas, que es también el 100% de los casos. Uno espera que la proporción de caras obtenidas en una serie de tiradas se aproxime a 0,5 (50%) cuando el número de tiradas es bastante alto.
La relación es la misma con respecto a la deriva genética. Cuanto mayor sea el número de individuos en la población, menor será la diferencia entre las frecuencias de una generación y la siguiente, aunque no debemos olvidar que lo que cuenta no es el número total de individuos en la población, sino su “tamaño eficaz”. El tamaño eficaz se define por aquellos individuos que dejan descendientes: la razón es que sólo los individuos reproductores transmiten sus genes a la generación siguiente, y si éstos son muy pocos, las consecuencias de la deriva pueden ser grandes (a efectos de la deriva genética, la existencia de pocos individuos reproductores, aunque la población total sea muy grande, es igual a una población muy pequeña). En general, para muchas clases de organismos se calcula que el tamaño efectivo de la población es aproximadamente un tercio del número total de individuos.
Los efectos de la deriva genética son normalmente pequeños de una generación a otra, dado que la mayoría de las poblaciones naturales constan de miles de individuos reproductores. Pero los efectos sobre muchas generaciones pueden ser importantes. Si no hubiera otros procesos de cambio evolutivo, tales como el de selección natural y el de mutación, las poblaciones llegarían al final a tener un solo alelo de cada gen. La razón es que tarde o temprano uno u otro alelo sería eliminado por la deriva genética. Debido a la mutación, los alelos desaparecidos de una población pueden reaparecer de nuevo y, si unimos a la ecuación la selección natural, podemos comprender que la deriva genética no tiene consecuencias importantes en la evolución de las especies salvo en aquellas que constan de poquísimos individuos.
Y este es precisamente el supuesto que Ernst Mayr llamó «efecto fundador». Variaciones fortuitas en las frecuencias alélicas similares a las debidas al efecto fundador tienen lugar cuando las poblaciones pasan a través de un «cuello de botella». Cuando el clima u otras condiciones son desfavorables, es posible que las poblaciones sufran una pérdida drástica de sus individuos y corran el riesgo de llegar a extinguirse. Sin embargo, si superan esa situación dramática, las poblaciones pueden recobrar su tamaño normal con el paso del tiempo aunque los genes portados por todos los miembros derivarán de los pocos genes presentes originalmente en los fundadores o supervivientes.
Representación gráfica de un cuello de botella poblacional (wikimedia commons).
En la historia evolutiva de nuestra especie se ha dado el caso de que las tribus eran diezmadas con frecuencia a causa de diversas enfermedades, bruscos cambios climáticos y otras circunstancias, aunque por suerte para nosotros, se recobraban a partir de los supervivientes o gracias a los inmigrantes de otras tribus. De esta forma, las diferencias entre las poblaciones actuales de seres humanos en la frecuencia de algunos genes parecen ser, al menos en arte, el resultado de los cuellos de botella por los que pasaron las poblaciones humanas primitivas.
Por otro lado tenemos la mutación. Podemos definir una mutación como cualquier cambio estable y heredable en la secuencia de nucleótidos del ADN. Cuando dicha mutación afecta a un sólo gen se denomina mutación génica; cuando es la estructura de uno o varios cromosomas la que se ve afectada, mutación cromosómica; y cuando una o varias mutaciones provocan alteraciones en todo el genoma se denominan mutaciones genómicas.
La tasa de mutación de un gen o una secuencia de ADN es la frecuencia en la que se producen nuevas mutaciones en ese gen o en la secuencia en cada generación. Una alta tasa de mutación implica un mayor potencial de adaptación en el caso de un cambio ambiental, pues permite a los individuos disponer de más variantes genéticas, lo que aumenta la probabilidad de que surja la variante adecuada necesaria para adaptarse al cambio ambiental. Del mismo modo, y como contrapartida, una alta tasa de mutación aumenta el número de mutaciones perjudiciales o deletéreas de los individuos y, por tanto, aumenta también la probabilidad de extinción de la especie.
Por lo tanto, una mutación en un lugar clave puede producir un cambio de características del organismo, que se presenta súbita y espontáneamente, y que se puede heredar o transmitir a la descendencia. Una vez en marcha, la selección natural actuará sobre ese cambio, manteniéndolo o eliminándolo en el caso de que permita o no al organismo adaptarse mejor al ambiente. También sucede en muchas ocasiones que una mutación no produce ningún efecto determinante.
Por último, el intercambio de genes entre poblaciones debido a la migración de los individuos entre poblaciones es otro factor importante de cambio genético. Si dos poblaciones difieren en las frecuencias de los alelos de algunos de sus genes, entonces el intercambio genético entre individuos de las poblaciones emigrante y local producirá un cambio de las frecuencias de los genes en cada una de ellas. Las migraciones humanas durante la expansión neolítica por ejemplo determinaron significativamente el tipo y la cantidad de variación genética de nuestra especie.
La sistemática o cómo clasificamos los organismos
Una vez que hemos analizado la teoría de la evolución y comprendido la enorme diversidad biológica que es capaz de generar la selección natural, podemos entender la importancia que tiene contar con un sistema que permita clasificar nuestro conocimiento del mundo animal, no sólo por necesidades intelectuales y de organización, sino porque un buen sistema de este tipo permite extraer importantes conclusiones evolutivas.
En biología, la sistemática es el estudio de la identificación, la taxonomía y la nomenclatura (estas últimas son disciplinas auxiliares que tratan del análisis de las características de un organismo con el propósito de clasificarlo y nombrarlo conforme a unas reglas establecidas). Así, la sistemática trata del estudio de la clasificación de las especies con arreglo a su historia evolutiva. Inicialmente su interés se centró en la observación de los distintos organismos, actividad que permitió una rápida acumulación de datos descriptivos, lo que a su vez llevó a mediados del siglo XVIII a la revolución de Carl von Linné, quien estableció normas precisas para nombrar y clasificar los seres vivos (para más información ver esta anotación). Por lo tanto, la sistemática se ocupa de la diversidad biológica tanto en un plano descriptivo como en el explicativo o interpretativo.
¿Y qué método se emplea para llevar a cabo esta clasificación? El campo que ahora nos interesa, el de la paleontología humana, vivió una revolución gracias a la cladística, un método de clasificación planteado por el entomólogo alemán Willi Henning en 1950 en su obra Grundzüge einer Theorie der phylogenetischen Systematik (Fundamentos de una teoría de la sistemática filogenética). Es un método de clasificación que solo utiliza líneas de descendencia en lugar de parecidos morfológicos para deducir parentescos evolutivos y que agrupa estrictamente a los organismos en función de su proximidad relativa a un antepasado común.
Expliquemos un poco mejor esta cuestión: lo que hace la cladística es agrupar a las especies en “clados”, que significa “un grupo, una rama del árbol filogenético” (o árbol evolutivo, los dos términos son prácticamente sinónimos). Para ello se utiliza como criterio de reconocimiento de pertenencia al grupo (o clado) la identificación de al menos un carácter apomorfo compartido por todos los miembros de ese grupo y heredado de su especie ancestral (una apomorfia es un carácter nuevo evolucionado a partir de otro preexistente. El carácter original y el derivado forman una pareja homóloga). Por tanto, la idea central es la llamada monofilia estricta, según la cual un grupo es monofilético si todos los organismos incluidos en él han evolucionado a partir de un antepasado común. De esta manera, un clado incluirá a un grupo de especies y a su antepasado común (el fundador del clado) a partir del cual se diversificaron ―al clado se le denomina también “grupo natural” (porque es un reflejo de la verdadera historia evolutiva) y “grupo monofilético” (porque tiene un solo origen).
Tipos de clados (wikimedia commons).
Para formar los clados a partir de las especies con las que se está trabajando, la cladística divide los caracteres en dos tipos. Unos son los caracteres primitivos, que se heredan de un antepasado anterior al nacimiento del clado y por eso se encuentran también en otros clados. Otros caracteres son los llamados derivados, que sólo se observan dentro del clado porque aparecieron con la especie fundadora del mismo. Son los caracteres derivados los únicos que cuentan en el análisis filogenético. Los caracteres “primitivos” se llaman en cladística “plesiomorfias”, y para los “derivados” se usa el término “apomorfia” como hemos indicado más arriba.
Para concluir con este apartado, decir que la cladogénesis hace referencia a la diversificación o multiplicación de linajes, de clados. Por ejemplo, la diversificación de los linajes que llevan al hombre, el chimpancé y el gorila, a partir de un linaje único que existía entre hace ocho y diez millones de años es un ejemplo de cladogénesis. Los ejemplos de cladogénesis son innumerables puesto que la evolución ha producido la extraordinaria diversidad del mundo viviente. Como términos sinónimos que podemos encontrarnos para referirse a la cladogénesis son “evolución divergente” y “evolución diversificadora”.
Con toda la información que hemos analizado sobre sistemática y clasificación, podemos comprender que la evolución de los seres vivos puede ser representada como un árbol, un árbol cuyas ramas se van dividiendo a medida que se alejan de la base del tronco. Esta base representa la especie ancestral común a todas las del árbol, y la ascensión hacia la copa representa el paso del tiempo. Sus ramas se corresponden a los linajes en evolución, algunos de los cuales eventualmente se extinguen, mientras otros llegan hasta el presente. Tales “árboles” se denominan “dendrogramas” (del griego dendron árbol), “árboles filogenéticos” o simplemente, “filogenias”:
Cladograma que representa la diversificación entre los gibones, orangutanes, gorilas, chimpancés y humanos.
Conclusiones
Espero que el concepto de evolución y los diferentes mecanismos por los que se produce se hayan entendido bien (en caso contrario, en los comentarios podemos incidir en los aspectos que sea necesario reforzar). Dicho esto, hay quienes sostienen que Darwin no estuvo muy acertado al elegir la palabra «selección», ya que este término parece indicar que existe en la naturaleza alguien que selecciona deliberadamente los organismos mejor adaptados. En realidad, como ya hemos explicado, los individuos «seleccionados» son sencillamente los que sobreviven después de que se hayan eliminado de la población los individuos peor adaptados o menos afortunados. Por eso se ha propuesto sustituir la palabra selección por la frase «eliminación no aleatoria». En definitiva, se usa la palabra «selección» para designar el conjunto de circunstancias responsables de la eliminación de algunos individuos.
Volvamos ahora a la imagen con la que iniciábamos esta anotación. Esta imagen es engañosa y contraproducente para comprender la evolución humana por dos motivos fundamentales. El primero guarda relación con la errónea idea de «dirección», de evolución hacia la complejidad y la «perfección» que nos transmite. Vemos una serie de pasos «evolutivos» que van desde el ser más pequeño y atrasado hacia el más erguido, musculoso y de tez clara que se representa al final. La idea que se quiere transmitir es obvia: el ser humano ha evolucionado de un animal pequeño y peludo para convertirse, con el paso del tiempo, en la cima de la naturaleza. Sin embargo, las cosas no han sucedido de forma tan aséptica; la evolución del hombre no ha sido una limpia carrera de relevos en la que, tras cinco cómodas etapas, ha surgido el ser humano moderno en la meta. Aunque lo explicaremos con más detalle en una anotación posterior, la evolución de nuestra especie se asemeja más a un arbusto que a un árbol. Los últimos avances en genética nos dicen que nuestro pasado evolutivo ha sido más intrincado, mucho más complejo de lo que nunca habíamos imaginado. Y algo muy importante, muchos de los linajes de nuestros antepasados se extinguieron sin más.
El segundo motivo por el que esta imagen no ha sido afortunada tiene que ver con la ya famosa frase «el hombre desciende del mono». El primer ser de la imagen tiene un parecido enorme con un chimpancé, y es un error muy común ―demasiado común― pensar que descendemos de ellos o de cualquier otro simio actual (gorilas u orangutanes).
Debemos tener claro que el hombre no desciende del chimpancé, ni del gorila, ni del mono aullador, ni del macaco japonés, sino de primates que hoy en día ya no existen. A la pregunta de si el hombre desciende del mono, la respuesta de un biólogo evolutivo sería un rotundo «no», el hombre no desciende del chimpancé (el término «mono» no es taxonómico y, en cualquier caso, nuestros antepasados son simios). Lo que sí está perfectamente documentado es que el hombre desciende de otras especies de homininos ya desaparecidos, y que éstos, a su vez, provienen de otros primates igualmente desaparecidos que dieron lugar a otras ramas evolutivas de las que descienden los chimpancés actuales. Tenemos que ser conscientes que también existen restos fósiles de los antepasados de chimpancés, gorilas, orangutanes etc.