Última actualizacón: 4 abril 2020 a las 18:38
En el grupo de los homininos arcaicos reunimos todos los taxones que no son inequívocamente Homo ni Paranthropus: incluye dos géneros, Australopithecus y Kenyanthropus. La paleoantropología tiene serios problemas para explicar cómo surgió el género Australopithecus. Hasta la fecha, la explicación más aceptada es que Ardipithecus dio lugar al nuevo género en el valle del Rift (otro de los motivos por los que el lugar donde fue hallado Sahelanthropus resulta tan inquietante). Así, apenas 200.000 años después de que Ar. Ramidus deambulara por las junglas de lo que hoy conocemos como Etiopía, un nuevo hominino hacía su aparición con rasgos inequívocos de un bipedismo obligado —aunque con posibles reminiscencias de locomoción arbórea—. Junto a la bipedia, el aumento del tamaño de los molares y del grosor del esmalte dental son las características básicas que definen a este grupo.
Los australopitecinos no constituyen en realidad un único clado ya que agrupa algunas pero no todas las especies descendientes de un antepasado común. Así, los científicos hacían una distinción informal entre ejemplares «gráciles» y «robustos» que ahora conocemos como dos géneros distintos: Australopithecus los primeros y Paranthropus los segundos. Por este motivo, hasta que no dispongamos de las muestras suficientes ni de los métodos de análisis adecuados para generar una filogenia fiable, tiene poco sentido revisar esta terminología que se encuentra muy extendida.

Yacimientos de los homininos arcaicos.
Australopithecus anamensis
Nombre taxonómico: Australopithecus anamensis ―Leakey, et al. (1995)― se halla restringido en la actualidad al este de África. El primer fósil descubierto fue la parte distal de un húmero izquierdo (KNM-KP 271) en 1967 en Kanapoi (Kenia). Otros yacimientos que han aportado fósiles al hipodigma son Allia Bay en Kenia, y el área de estudio del Middle Awash en Etiopía. Antigüedad: entre 4,19 y 3,72 Ma.

Holotipo Australopithecus anamensis.
KNM-KP 29281, mandíbula en vista oclusal (superior izquierda), lateral derecha (superior derecha) y medial (centro a la derecha). En la parte inferior, el temporal en vista inferior (abajo a la izquierda) y lateral (abajo a la derecha). (Leakey, et al. 1995)
Espécimen tipo: KNM-KP 29281. Se trata de una mandíbula con la dentición completa y un fragmento del hueso temporal que pertenece probablemente al mismo individuo. Ambos fueron recuperados en 1994 en Kanapoi.
Características y comportamiento: Entre los fósiles contamos con mandíbulas, dientes y elementos postcraneales de las extremidades superiores e inferiores. La mayoría de las diferencias entre Au. anamensis y Au. afarensis guardan relación con la dentición. En algunos aspectos, los dientes de Au. anamensis son más primitivos que los de Au. afarensis (con unos caninos grandes), pero en otros guardan parecido con los parántropos (esmalte dental grueso). Esta especie aporta la primera evidencia de incremento en el tamaño de los molares, que se asocia con el consumo de alimentos más duros y su vida en la sabana.
Los restos de las extremidades superiores son similares a los de Au. afarensis (con una morfología similar a la de los primates arbóreos), pero la tibia tiene rasgos asociados con la bipedia.
En 2016 se recuperó el primer cráneo asociado a especie (catalogado como MRD-VP-1/1). Tras dos años de trabajos se publicaron dos estudios, uno sobre la descripción del fósil y otro sobre el contexto geológico y la edad del mismo. La datación del fósil en 3,8 Ma nos indica que Australopithecus afarensis convivió en el tiempo con Australopithecus anamesis como mínimo durante 100.000 años. Esto pone en tela de juicio la hipótesis manejada hasta ahora de que Au. afarensis fuera el descendiente directo de Au. anamensis.
Referencia
- Leakey, M. G., et al. (1995), «New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya«. Nature, vol. 376, núm. 6541, p. 565-571.
Australopithecus afarensis
Nombre taxonómico: Australopithecus afarensis ―Johanson, et al. (1978)―. Se conoce únicamente en yacimientos del este de África. El primer fósil de esta especie, un fragmento de maxilar (Garusi 1) se recuperó en 1939 en Laetoli (Tanzania). La mayor parte del hipodigma proviene de Hadar pero otros yacimientos de Etiopía y Kenia han aportado más restos. Antigüedad: entre los 3,6 y 3 Ma (cerca de 4 Ma si se confirma su presencia en Belohdelie y Fejej).

Holotipo Australopithecus afarensis.
LH 4, vista oclusal. (Johanson, et al. 1978)
Espécimen tipo: LH 4. El espécimen tipo es una mandíbula hallada en Laetoli (Tanzania) en 1974 por el equipo de Mary Leakey ―Leakey, et al. (1976)―.
Características y comportamiento: Este es el hominino más antiguo del que contamos con un registro fósil bastante completo. El hipodigma incluye un cráneo bien conservado, cráneos parciales y fragmentados, muchas mandíbulas inferiores así como suficientes huesos de las extremidades como para poder estimar la estatura y masa corporal. Quizás el espécimen más conocido sea AL-288, la famosa «Lucy», una hembra adulta de la que conservamos casi el 80% de su esqueleto (si tomamos en cuenta la simetría bilateral). La mayoría de las estimaciones de la masa corporal la sitúan en el rango de los 30 a 45 kg de peso, una altura entre 1,10 y 1,30 metros de alto, y con una capacidad craneal estimada entre 400 y 550 cm3.
Tiene unos incisivos más pequeños que los de los actuales chimpancés, pero sus premolares y molares son relativamente más grandes. La comparación de los restos sugiere que las extremidades inferiores de AL-288 son sustancialmente más cortas que las de los humanos modernos de una estatura parecida. Tanto la forma de la pelvis como estas extremidades cortas sugieren que, aunque Au. afarensis era capaz de andar erguido, no estaba adaptado para un bipedismo prolongado. Tendría por tanto una locomoción particular: bípeda pero manteniendo cierta capacidad de desplazamiento por los árboles. Estas pruebas indirectas de su locomoción se completan con el descubrimiento de las «huellas de Laetoli», atribuidas a miembros de esta especie.
Las extremidades superiores, sobre todo la mano y la cintura escapular, poseen una morfología que se asocia a la locomoción arbórea. Aunque un estudio reciente argumenta que el dimorfismo sexual en este taxón está escasamente desarrollado, la mayoría de los investigadores opinan lo contrario.
Referencias
- Johanson, D. C.; White, T. D. y Coppens, Y. (1978), «A new species of the genus Australopithecus (primates: Hominidae) from the Pliocene of eastern Africa«. Kirtlandia, núm. 28, p. 2-14.
- Leakey, M. D., et al. (1976), «Fossil hominids from the Laetolil Beds«. Nature, vol. 262, núm. 5568, p. 460-466.
Australopithecus deyiremeda
Nombre taxonómico: Australopithecus deyiremeda ―Haile-Selassie, et al. (2015)― Recuperado en el área de estudio de Woranso-Mille, en la región de Afar (Etiopía). Antigüedad: entre 3,5 y 3,3 Ma.

Holotipo Australopithecus deyiremeda.
BRT-VP-3/1. a) vista oclusal; b) vista lateral; c) vista superior; d) vista medial; e) vista anterior. (Haile-Selassie, et al. 2015)
Espécimen tipo: BRT-VP-3/1. Un maxilar izquierdo con un incisivo y un molar.
Características y comportamiento: Pese a que cronológica y geográficamente coincide con Au. afarensis, los descubridores defienden que se trata de una nueva especie dadas las diferencias en la morfología de la mandíbula y el maxilar. Aún hay pocos restos para poder ofrecer un mayor detalle.
Referencia
- Haile-Selassie, Y., et al. (2015), «New species from Ethiopia further expands Middle Pliocene hominin diversity«. Nature, vol. 521, núm. 7553, p. 483-488.
Kenyanthropus (Australopithecus) platyops
Nombre taxonómico: Kenyanthropus platyops ―Leakey, et al. (2001)―. El primer fósil descubierto (KNM-WT 38350) era un fragmento de maxilar izquierdo recuperado en 1998 en Lomekwi, Turkana occidental (Kenia) por el equipo de Meave Leakey. Antigüedad: entre 3,5 y 3,3 Ma.

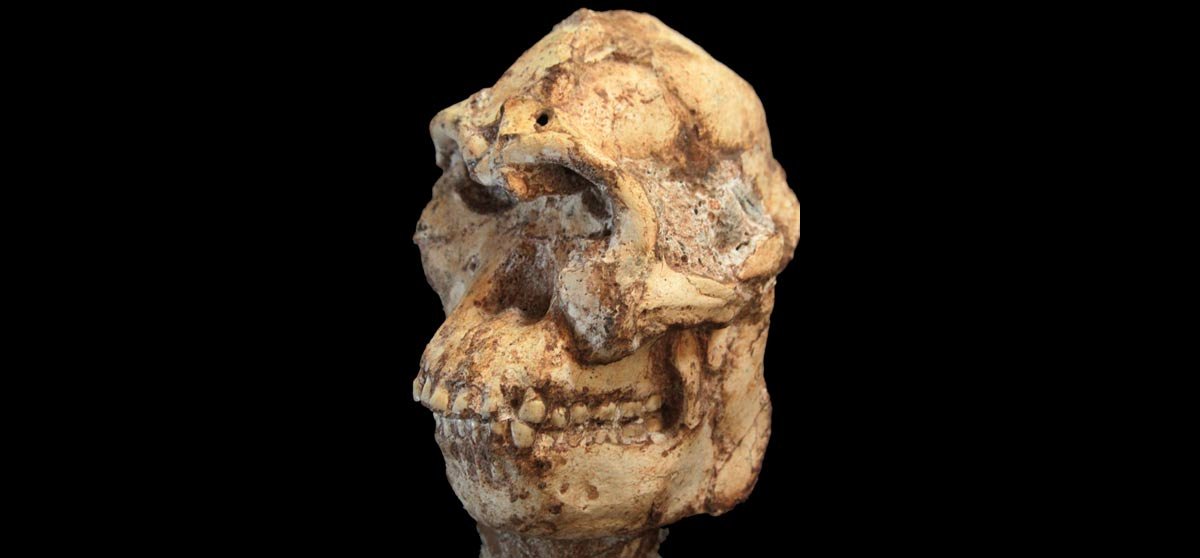
Holotipo Kenyanthropus platyops.
KNM-WT 40000. a) Vista lateral (la línea marca la separación entre el neurocráneo bastante distorsionado y la cara bien conservada; b) Vista superior; c) Vista anterior; d) Vista oclusal del paladar. (Leakey, et al. 2001)
Espécimen tipo: KNM-WT 40000. Es un cráneo relativamente completo pero deformado hallado en 1999 en Lomekwi.
Características y comportamiento: El cráneo tiene una configuración «grácil» según los autores del hallazgo, pero su antigüedad era mayor que la de cualquier otro espécimen del linaje grácil. Las principales razones que llevaron a Leakey y colaboradores a no asignar este material a Au. afarensis fueron dos: primero, el rasgo derivado que más salta a la vista, el de un plano muy vertical por debajo de la nariz que le proporciona el aspecto de cara plana al que se refiere su nombre; y en segundo lugar, unos molares relativamente pequeños pero con un esmalte dental parecido a Au. afarensis. A pesar de esta combinación única de morfología facial y dental, algunos autores como Tim White sostienen que un nuevo taxón no está justificado porque el cráneo puede ser el cráneo deformado de un Au. afarensis. Además, como hemos señalado al inicio, la estrategia adaptativa distintiva que podría deducirse de esa cara plana —y que sería necesaria para definir un nuevo género— no queda nada clara (de ahí que la mayoría de especialistas se refieran a este ejemplar como Australopithecus platyops).
Referencias
- Leakey, M. G., et al. (2001), «New hominin genus from eastern Africa shows diverse middle Pliocene lineages«. Nature, vol. 410, núm. 6827, p. 433-440.
- White, T. (2003), «Early Hominids – diversity or distortion?«. Science, vol. 299, núm. 5615, p. 1994-1997.
Australopithecus bahrelghazali
Nombre taxonómico: Australopithecus bahrelghazali ―Brunet, et al. (1996)―. Se ha identificado únicamente por una mandíbula parcial y un diente aislado encontrados en la región de Bahr el Ghazal en del desierto del Djourab (República de Chad) 1. Antigüedad: entre 3,5 y 3 Ma.

Holotipo Australopithecus bahrelghazali. KT 12/H1, a) Vista oclusal; b) vista anterior; c) vista vestibular derecha. (Brunet, et al. 1996).
Espécimen tipo: KT 12/H1. Se trata del fragmento de una mandíbula de un adulto que contiene un incisivo, los dos caninos y los dos premolares de ambos lados.
Características y comportamiento: Los descubridores mantienen que el esmalte dental más grueso lo distingue de Ar. ramidus, mientras que una sínfisis mandibular más pequeña y vertical (indicativa de una cara con un menor prognatismo) lo distinguen de Au. afarensis. De hecho, en un principio, la clasificación provisional de esta mandíbula fue la de Australopithecus aff. afarensis, pero un año más tarde, cuando Brunet lo comparó con los restos de Au. afarensis conservados en el Museo Nacional de Addis Abeba, se revisó y les otorgó el rango de nueva especie.
Parece una variante regional de Au. afarensis —y de hecho muchos investigadores agrupan estos restos en esa especie— pero el descubrimiento de Chad extiende sustancialmente el alcance geográfico de los primeros homininos y nos recuerda que los hitos importantes en la evolución humana (como la especiación y la extinción) pueden haber ocurrido bien lejos de las pequeñas regiones (en comparación con el tamaño del continente africano) donde existen yacimientos de los primeros homininos.
Referencia:
- Brunet, M., et al. (1996), «Australopithecus bahrelghazali, une nouvelle espèce d’Hominidé ancien de la région de Koro Toro (Tchad)«. Comptes rendus de l’Académie des sciences. Série 2. Sciences de la terre et des planètes, vol. 322, núm. 10, p. 907-913.
Australopithecus africanus
Nombre taxonómico: Australopithecus africanus ―Dart (1925)―. La mayoría de los fósiles de este espécimen provienen de dos cuevas, Sterkfontein y Makapansgat, aunque también se han recuperado en la cueva de Gladysvale. Antigüedad: entre 3 y 2,4 Ma.

Holotipo Australopithecus africanus. Taung 1, en vistas frontal, lateral derecha e inferior. (Dart, 1925)
Espécimen tipo: Taung 1. Se trata de un cráneo juvenil con un molde endocraneal parcial, recuperado en 1924 en Taung, Sudáfrica. Curiosamente, se pone en duda el verdadero sentido evolutivo del «niño de Taung» ya que la enorme cantidad de ejemplares procedentes de los otros yacimientos presentan rasgos bien distintos a éste. Estas dudas se ven incrementadas cuando tenemos en cuenta que Taung 1 es el único espécimen de hominino hallado en ese yacimiento.
Características y comportamiento: Con Au. africanus contamos con uno de los mejores registros fósiles de los homininos antiguos. Disponemos de abundantes muestras del cráneo, la mandíbula y la dentición. Por otro lado, el esqueleto postcraneal –y particularmente el esqueleto axial (la parte central del cuerpo formada por el torax, columna vertebral, cráneo, pelvis, etc.)– está peor representado, pero al menos tenemos una muestra de cada uno de los huesos largos, aunque gran parte de los fósiles han sido aplastados por rocas antes de que fosilizaran y están deformados.
La imagen que surge de los análisis morfológicos y funcionales sugiere que aunque Au. africanus era capaz de andar erguido (era bípedo) estaba mejor adaptado para desplazarse por los árboles (era un bípedo ocasional y no obligado) que otros taxones arcaicos como Au. afarensis. Tenía unos premolares relativamente grandes, y salvando los caninos reducidos, el cráneo es de aspecto simiesco. Su capacidad craneal media es de cerca de 485 cm3. Los restos hallados en Sterkfontein sugieren que tanto las hembras como los machos presentan diferencias en su tamaño corporal –dimorfismo sexual– pero no en el grado de Au. afarensis.
Referencia
- Dart, R. A. (1925), «Australopithecus africanus: the man-ape of South Africa«. Nature, vol. 115, núm. 2884, p. 195-199.
Australopithecus sediba
Nombre taxonómico: Australopithecus sediba —Berger, et al. (2010)—. Todos los fósiles de este taxón se han recuperado en un único yacimiento: Malapa (Sudáfrica). Estudios radiométricos combinados con paleomagnetismo han proporcionado una fecha consistente para este ejemplar de 1,97 Ma.

Holotipo Australopithecus sediba.
UW88-50 (MH1) elementos craneodentales en A. Vista superior; B. Vista frontal; C. Vistas laterales y D. mandíbula juvenil UW88-8 (MH1)
(Berger, et al. 2010)
Espécimen tipo: MH 1. Un ejemplar compuesto por un cráneo juvenil (UW88-50), una mandíbula fragmentada (UW88-8) y diversas partes del esqueleto.
Características y comportamiento: Se distingue de Au. anamensis, Au. afarensis y Au. garhi por la falta de las apomorfias típicas de esos australopitecos, como la proyección de la cara (prognatismo). Tampoco posee un gran aparato mandibular, la megadontia extrema de premolares y molares y las marcas pronunciadas de inserción muscular en el cráneo. En definitiva, estamos ante un aparato masticador menos desarrollado, con una cara y mandíbula de tamaño reducido.
Según sus descubridores Au. sediba se acerca más a Au. africanus, sobre todo por su «gracilidad» y la baja capacidad craneal. Sin embargo, las diferencias también son importantes y hace que esta especie se parezca a los ejemplares antiguos de Homo. El análisis de los fósiles del pie desvela un conjunto de rasgos primitivos asociados a la capacidad de trepar por los árboles, así como otros derivados que apuntan a la bipedia. Lo mismo sucede con los huesos de la mano y la muñeca, que indican una capacidad para desplazarse por los árboles y un agarre de precisión que podría indicar el uso de herramientas.
Referencia
- Berger, L. R., et al. (2010), «Australopithecus sediba: a new species of Homo-like australopith from South Africa«. Science, vol. 328, núm. 5975, p. 195-204.
![]()
Notas
- El mismo yacimiento donde años más tarde se descubrió a Sahelanthropus tchadensis. ↩